CONCEPTES BÀSICS
D'EVOLUCIÓ
Lluis Serra i Camo - curs 1992-1993
Evolució significa canvis en els éssers
vius durant llargs períodes de temps. L'evolució per selecció
natural es pot comprovar científicament. És l'única
teoria unificadora de la Biologia. Tal i com Dobzhansky indicava, "rés
te sentit en Biologia sinó és sota l'aspecte evolutiu".
Les modificacions evolutives dels éssers
vius depenen de canvis ambientals externs i de mutacions pre-adaptatives
innovadores. En qualsevol moment, la forma del futur canvi no es pot predir,
a no ser condicionalment.
Evolució significa canvi en la morfologia
i el comportament dels organismes entre generacions (cal explicitar "entre
generacions" per no incloure en la definició aquells canvis del
desenvolupament, que tenen lloc durant la vida d'un organisme). A més,
l'evolució de la vida s'ha donat seguint un model dicotòmic,
un procés de ramificació. La gran varietat d'espècies
actuals s'ha generat per un procés repetit de separació de
llinatges, a partir d'un únic avantpassat comú.
El procés de canvi i la separació
de llinatges són, doncs, els mecanismes fonamentals de l'evolució.
L'adaptació és un altre concepte evolutiu fonamental. Fa
referència a aquelles propietats dels éssers vius que els
permeten sobreviure i reproduir-se a la natura. El camuflament n'és
un exemple. Les espècies camuflades tenen models de coloració
i detalls morfològics i de comportament que les fan menys visibles
en els seus habitats naturals. El camuflament ajuda a l'organisme a sobreviure
fent-lo menys visible als seus enemics naturals. El camuflament és,
doncs, adaptatiu. En la teoria de Darwin, l'adaptació s'explica
per selecció natural.
La selecció natural significa que alguns
individus en una població produeixen més descendents que
els altres. Com a conseqüència d'això, les característiques
genètiques d'aquests individus incrementaran en freqüència
a la població al llarg del temps; la composició genètica
de la població canviarà, així, automàticament.
HISTÒRIA DE LA BIOLOGIA EVOLUTIVA
-
Idees evolutives abans de Darwin
-
Teoria de Darwin (1859)
-
L'eclipsi de Darwin
-
La síntesi moderna (1920 - 1950)
-
Desenvolupaments posteriors (1960 - ...)
1 - Idees evolutives abans de Darwin
El caràcter fix de les espècies
era l'opinió ortodoxa en els temps de Darwin, encara que alguns
autors ja havien manifestat el seu desacord.
J. B. Lamarck (1744 - 1829)
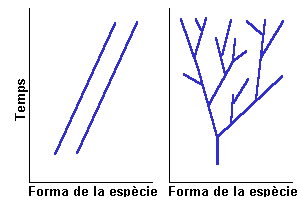 Les
espècies es transformen al llarg del temps en altres espècies.
La figura 1 representa la concepció lamarckista de l'evolució
i com es diferencia del concepte darwinista. Lamarck suposava que els llinatges
de les espècies no s'extingien, sinó que canviaven d'una
forma a un altre (anagènesi); el llinatges no es ramificaven. Va
proposar un mecanisme que explicava el canvi anagenètic: l'herència
dels caràcters adquirits o la teoria de l'ús i el desús.
Suposa que les mutacions són post-adaptatives, la qual cosa s'ha
rebutjat a nivell experimental (per exemple la prova de la fluctuació
de Delbrück i Luria, o la resistència de Drosophila
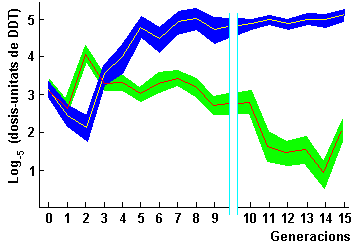
al DDT). A la figura 2 s'hi representa esquemàticament l'experiment
de la resistència al DDT, i a la figura 3 els resultats de la selecció.
Les
espècies es transformen al llarg del temps en altres espècies.
La figura 1 representa la concepció lamarckista de l'evolució
i com es diferencia del concepte darwinista. Lamarck suposava que els llinatges
de les espècies no s'extingien, sinó que canviaven d'una
forma a un altre (anagènesi); el llinatges no es ramificaven. Va
proposar un mecanisme que explicava el canvi anagenètic: l'herència
dels caràcters adquirits o la teoria de l'ús i el desús.
Suposa que les mutacions són post-adaptatives, la qual cosa s'ha
rebutjat a nivell experimental (per exemple la prova de la fluctuació
de Delbrück i Luria, o la resistència de Drosophila
al DDT). A la figura 2 s'hi representa esquemàticament l'experiment
de la resistència al DDT, i a la figura 3 els resultats de la selecció.
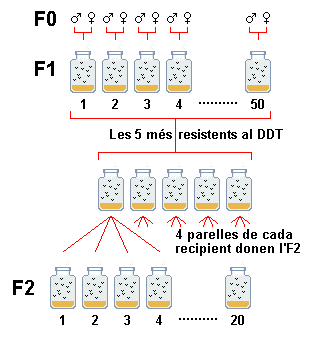 |
S'aïllen 50 parelles d'una soca en 50 ampolles.
Cada parella dona lloc a una germandat. Cada germandat s'analitza per la
resistència al DDT: es divideix en 4 subpoblacions de les quals
3 es sotmeten a concentracions diferents de DDT - per exemple 1/25, 1 i
25 micrograms per ml. Així es pot determinar la dosi letal 50, que
causa la mort de la meitat de la població. La quarta subpoblació,
que no ha estat mai en contacte amb el DDT, és el testimoni. S'esculleixen
les 5 germandats més resistents i les 5 menys resistents. La 4ª
subpoblació de cadascuna (que no ha estat en contacte amb el DDT)
s'utilitza per fundar la generació següent: s'agafen 4 parelles
de cadascuna de les 5 germandats, en total 20 parelles. |
| Fig a |
|
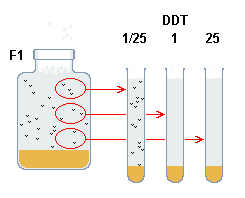 |
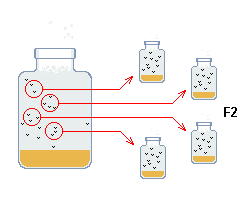 |
| Fig b: Determinació de la resistència
al DDT |
Fig c: Formació de la següent generació |
Cadascuna s'analitza de la mateixa manera a les generacions
següents. Després de 14 generacions de selecció, la
soca seleccionada per millor resistència era 625 vegades més
resistent que l'original.
En part per reacció contra Lamarck, Cuvier
i la seva escola van aconseguir que la major part de biòlegs acceptessin
la teoria del caràcter fix de les espècies. Cuvier, però,
considerava que les espècies es podien extingir, i les seves anàlisis
del registre fòssil el portaren a admetre la teoria del catastrofisme.
2 - Teoria de Darwin (1859)
L'observació de la variació geogràfica
va ser, probablement, la causa que va portar a Darwin a admetre que les
espècies podien canviar. El següent pas consistia en inventar
una teoria que expliqués perquè canviaven les espècies.
Darwin va trobar l'explicació llegint el llibre de Malthus "Essay
on Population". Degut a la lluita per la supervivència, els
organismes o formes més adaptades produiran més descendents
i automàticament incrementarà la seva freqüència
a la població. A mesura que aquest procés continuï,
en últim terme es produiran noves espècies.
Una de les principals dificultats de la teoria
era que no es coneixien els mecanismes de l'herència, i per tant
tampoc l'origen de la variabilitat genètica a les poblacions naturals.
3 - L'eclipsi de Darwin
Els primers mendelians, com Hugo de Vries i William
Bateson, s'oposaren a la teoria de Darwin de la selecció natural.
Aquests autors estudiaven l'herència de caràcters qualitatius,
amb una influència molt forta sobre el fenotip i generalitzaren
els seus resultats al procés evolutiu. Per ells, l'evolució
es verificava a grans salts, mitjançant macromutacions. Ara bé,
el mendelisme no era acceptat universalment en el període 1900 -
1920: l'escola biomètrica, encapçalada per Karl Pearson,
estudiava diferències qualitatives entre els individus i desenvolupà
uns mètodes estadístics per descriure la transmissió
de les distribucions dels caràcters entre generacions. Aquesta escola
considerava l'evolució com un canvi gradual i no pas com un conjunt
de macromutacions.
4 - La síntesi moderna (1920 - 1950)
Sir Ronald Aymler Fisher va demostrar que els
resultats obtinguts per l'escola biomètrica es podien explicar segons
els principis mendelians. El següent pas consistia en demostrar que
la selecció natural era compatible amb l'herència mendeliana.
El treball teòric va ser fet principalment per R. A. Fisher, J.
B. S. Haldane i Sewall Wright. La síntesi de la teoria de Darwin
de la selecció natural amb la teoria mendeliana es coneix com el
Neo-Darwinisme,
la Teoria sintètica de l'evolució o La nova síntesi,
d'acord amb el títol del llibre de Julian Huxley: "Evolution: the
Modern Synthesis".
El Neodarwinisme inclou 5 principis bàsics:
-
Les poblacions naturals contenen una gran quantitat
de variabilitat genètica, produïda per mutació preadaptativa
i recombinació, i no pas per respostes postadaptatives dirigides
per les necessitats dels organismes.
-
L'evolució de les poblacions està determinada
per la selecció natural, així com també per el flux
genètic i la deriva genètica, i es caracteritza pel canvi
de les frequències gèniques.
-
La variabilitat genètica adaptativa produeix
petits canvis graduals del fenotip, que s'acumulen en els llinatges evolutius
durant llargs períodes de temps.
-
Les poblacions aïllades geogràficament
tendeixen a divergir i aquest procés de divergència te com
a resultat l'especiació, o formació de grups aïllats
reproductivament.
-
L'acumulació gradual de diferències
genètiques, mitjançant l'acció de la selecció
natural, determina també l'aparició de les categories taxonòmiques
per sobre del nivell d'espècie.
5 - Desenvolupaments posteriors (1960 - ...)
a) Seleccionisme i neutralisme.
A l'any 1968, Motoo Kimura va proposar l'anomenada
Teoria
neutralista de l'evolució molecular: la major par dels canvis
evolutius a nivell molecular no estàn determinats per la selecció
natural sinó per la deriva genètica de mutacions neutres.
La teoria de Kimura no solament s'ajustava a les dades de que es disposaven
sobre les seqüències d'aminoàcids de les proteïnes,
sinó també, a partir de 1980, a les dades sobre seqüènis
nucleotídiques. Les seqüències de DNA han sigut també
importants en un altre aspecte: han demostrat que el genoma té una
estreuctura particular, amb clusters de gens i regions no codificadores.
Cadascuna d'aquestes regions del genoma té la seva pròpia
forma d'evolucionar. La teoria neutralista no sugereix, però, que
la deriva explica tots els canvis evolutius; encara es necessita la selecció
natural per explicar les adaptacions. Es possible, però, que les
adaptacions que s'observen en els organismes es produèixin per una
minoria de canvis evolutius en el DNA. La teoria neutralista afirma que
l'evolució a nivell del DNA i les proteïnes (no les adaptacions)
depén sobre tot de processos aleatoris. La major part dels canvis
evolutius a nivell molecular serien no adaptatius.
b) A vegades s'ha dit que la selecció natural
produeix adaptacions pel be de l'espècie o pel be del grup. De fet,
la selecció natural produeix adaptacions en benefici de l'individu
i això pot tenir coseqüències deletèries per
el grup o l'espècie. La funció del sexe n'és un exemple.
 c)
Puntualisme
i Gradualisme.
c)
Puntualisme
i Gradualisme.
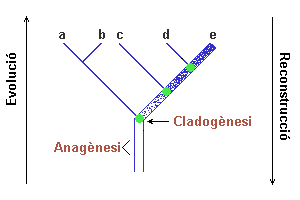
Anagènesi: L'espècie canvia
al llarg del temps fins que es tant diferent de l'espècie progenitora
que es classifica com una nova espècie. (figura 4)
Cladogènesi: L'especiació te
lloc com un procés de ramificació en el que una espècie
es converteix en dues. (figura 4)
Tres teoríes de l'història de la vida:
(figura 5)
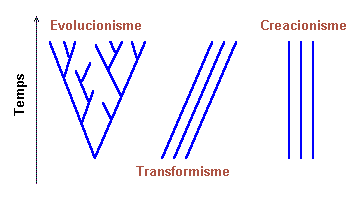
-
Evolucionisme. Les espècies tenen un avantpassat
comú i es transformen.
-
Transformisme. Les espècies tenen orígens
diferents i es transformen.
-
Creacionisme. Les espècies tenen orígens
diferents i no es transformen.
El registre fòssil planteja un problema al
darwinisme: no presenta transicions evolutives graduals i contínues.
Sovint una espècie apareix de manera sobtada, persisteix durant
un cert príode i s'extingeix. Aleshores es pot trobar un altre espècie,
però sense formes de transició. Darwin va explicar aquestes
discontinuïtats argumentant que el registre fòssil era incomplet.
(figura 5)
1972. Eldredge i Gould: Els paleontòlegs
no han interpretat bé el neodarwinisme, han buscat exemples d'especiació
filètica segons la qual les noves espècies s'originen per
transformació i no pas per cladogènesis d'una línea,
i han interpretat totes les observacions en funció d'aquest model
(figura 6). 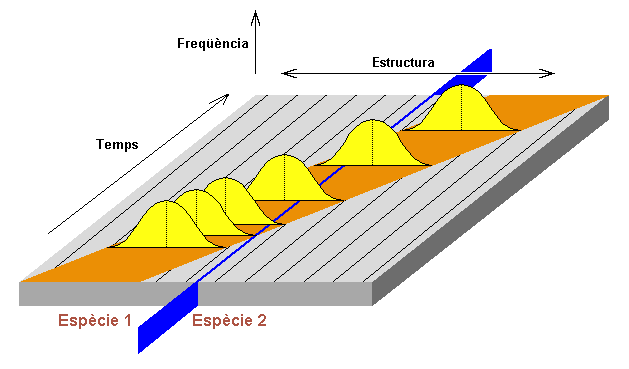
Aquest model, anomenat Gradualisme filètic
considera el següent:
-
Les noves espècies s'originen per transformació
d'una població ancestral que dóna lloc a descendents amb
modificacions al llarg de les generacions.
-
La transformació és lenta i contínua.
-
La transformació afecta a una gran nombre
d'individus, en general a tota la població.
-
La transformació té lloc en tota, o
en una gran part, de la distribució geogràfica de l'espècie.
Eldredge i Gould utilitzen el model de la població
perifèrica, aïllada, per demostrar que el registre fòssil
pot reflectir un mecanisme diferent al gradualisme filètic. Si les
noves espècies s'originen al.lopàtricament, a partir de poblacions
petites i aïllades, aleshores, el registre fòssil en un lloc
podria no reflectir el fenòmen de l'especiació. Podrien no
formar-se fòssils a la regió perifèrica, aïllada
on ha tingut lloc l'especiació; a més, la grandària
petita de la població aïllada fa que sigui poc probable que
es conservin fòssils de les formes intermèdies. Si, posteriorment,
la nova espècie envaeix l'àrea ocupada per l'espècie
ancestral (la qual cosa podria succeir si les dues espècies competissin
o fossin prou diferents per coexistir ecològicament), es podrien
trobar fòssils de les dues espècies en el mateix lloc, sense
formes de transició. Eldredge i Gould resumeixen així la
seva teoria de l'equilibri puntuat:
-
Les noves espècies s'originen per desdoblament
de llinatges.
-
L'especiació és ràpida.
-
Una petita subpoblació de la forma ancestral
dona lloc a la nova espècie.
-
Les noves espècies s'originen en una petita
part de la distribució geogràfica de l'espècie ancestral,
en una zona aïllada i perifèrica.
La diferència fonamental entre la teoria del
gradualisme filètic i de l'equilibri puntuat fa referència
a les taxes d'evolució durant i entre els fenòmens d'especiació.
Segons el gradualisme filètic, les taxes són semblants en
amdós moments. Segons la teoria de l'equilibri puntuat, la taxa
és molt més ràpida en el moment de l'especiació;
entre les puntuacions hi han llargs períodes d'estasi (sense canvis,
o molt pocs canvis) (figura 6). El gradualisme filètic admet l'especiació,
però difereix de l'equilibri puntuat per el fet d'admetre les mateixes
taxes evolutives durant i entre els fenòmens d'especiació.
ESPECIACIÓ
Un dels grans problemes de la Biologia consisteix
en explicar com un procés d'evolució contínua pot
originar els grups morfològicament discontinus que anomenem espècies.
En relació a l'especiació ens podríem plantejar una
sèrie de qüestion fonamentals.
-
Quants gens han de canviar per obtenir una nova espècie?.
-
Aquests gens, es troben en loci concrets dels cromosomes?;
hi han cromosomes que el seu contingut genètic sigui especialment
important per el fenòmen de l'especiació?.
-
Els canvis convencionals de les freqüències
gèniques són els únics importants en el procés
d'especiació o són necessaris altres mecanismes evolutius?.
-
Quina és l'importància relativa de
la selecció i la deriva genètica?.
-
Quina importància té l'aïllament
geogràfic?.
La major part de biòlegs evolutius accepten
el concepte biològic d'espècie proposat per Mayr: grups de
poblacions d'organismes que poden encreuar-se i donar descendència
fèrtil, aïllats reproductivament d'altres grups. Les barreres
reproductives que separen els membres de les diferents espècies
es dividèixen en mecanismes d'aïllament prezigòtics
(discriminació de la parella, diferències d'hàbitat,
polinització per diferents insectes...) i postzigòtics (inviabilitat,
esterilitat i degradació dels híbrids). L'aïllament
reproductor, conjuntament amb la selecció i la deriva genètica
han contribuit en crear i extendre les diferències morfològiques
que existèixen entre les espècies que ocupen la mateixa àrea.
Per tant, entendre l'origen dels mecanismes d'aïllament és
entendre l'origen de les espècies.
Segons els neodarwinistes, per entendre l'origen
d'aquests mecanismes d'aïllament cal suposar que existèixen
barreres geogràfiques entre les poblacions. Aquest aïllament
físic o al.lopatria, porta inevitablament a la divergència
evolutiva mitjançant la selecció natural i/o la deriva genètica.
Després d'un temps suficient, l'aïllament reproductor es desenvolupa
com una conseqüència més d'aquesta diferenciació
genètica.
Encara que pot assolir un aïllament reproductor
total durant el període de diferenciació geogràfica,
s'ha suggerit sovint que la selecció natural actuant sobre poblacions
no aïllades completament que han esdevingut simpàtriques (ocupen
la mateixa àrea), podria completar l'especiació mitjançant
la fixació d'aquells al.lels que redueixen l'aparellament interespecific,
maximitzant per tant l'eficacia biològica de la descendència.
Aquest procés s'anomena reforçament. Encara que algunes
dades experimentals semblen recolçar aquest fet, els models teòrics
demostren que és poc probable. L'importància de l'al.lopatria
és potser la única "regla de l'especiació" ampliament
acceptada.
Malgrat tot, cada vegada s'admet més, també,
que l'aïllament geogràfic no és sempre necessari, i
que es pot donar l'especiació simpàtrica dintre d'una
població. La probabilitat d'aquest fet és un dels aspectes
més discutits de les teories de l'especiació, però
alguns models i experiments demostren que es pot desenvolupar un aïllament
reproductor entre individus que poden estar potencialment en contacte.
Un estudi recient demostra que les etapes inicials de l'especiació
simpàtrida, - polimorfisme de nínxol, amb una reducció
del flux genètic -, poden donar-se a la natura: la diferenciació
genètica existent entre "races d'hoste" de la mosca tefrítida
nord-americana Rhagoletis pomonella així ho demostra. L'especiació
simpàtrica sembla estar limitada a tipus especials d'organismes,
com els paràsits i parasitoides fitòfags i zoòfags.
Es curiós que pràcticament no hi
han proves experimentals que demostrin l'origen de l'aïllament reproductor
com a conseqüència de la selecció o de la deriva, en
poblacions al.lopàtriques. Existeix un exemple a la natura d'un
cas d'aïllament reproductor com a conseqüència d'una adaptació:
els gens que determinen tolerància al coure de les poblacions de
plantes de l'espècie Mimulus guttatus que viuen al voltant de les
mines, són letals en els híbrids interpoblacionals.
Altres formes d'especiació no geogràfica
són l'especiació parapàtrica i l'estasipàtrica:
Especiació parapàtrica:
també anomenada clinal; diferenciació local d'una espècie
dintre d'una clina geogràfica contínua, superant la força
cohesiva del flux genètic produit per migració.
Especiació estasipàtrica:
aparició d'una reordenació cromosòmica, que s'origina
en algún lloc de l'àrea ocupada per l'espècie i que
redueix l'eficàcia biològica en heterozigosi.
Generalment l'aïllament reproductor està
controlat per més d'un gen: Dobzhansky i Muller van proposar un
model senzill de 2 loci; Tenim una espècie ancestral de genotip
A1A1B1B1 que evoluciona cap a A1A1B2B2 en una població i cap a A2A2B1B1
en un altre. S'obtindria una especiació si la interacció
epistàtica entre els al.lels A2 i B2 determinés esterilitat
completa o inviabilitat en els híbrids.
A Drosophila l'aïllament reproductor
té sovint una base poligènica amb un mínim de 5 a
9 loci. En els cargols l'aïllament pre-zigòtic pot venir determinat
per una mutació que canvia la direcció de cargolament de
la closca, impedint l'aparellament entre el normal i el mutant. La fixació
de l'al.lel mutant depèn del tipus d'herència d'aquest gen
(efecte matern) i de la poca movilitat dels cargols.
Es conèixen altres casos en els que un
mecanisme d'aïllament depén de tres o menys loci: per exemple,
la inviabilitat dels híbrids entre varietats de "platys" (peixos
d'aigua dolça), determinada per la formació de tumors (hi
intervenen 2 gens): els gens que determinen l'aparició de taques
de pigment en els Platys, ocasionen melanomes mortals en els híbrids
amb la varietat "swordtail"; aquesta anomalia depèn d'un oncogen
lligat al cromosoma X que codifica a un receptor de la tirosina-quinasa,
el qual està suprimit normalment en els Platys per un al.lel
dominant autosòmic. La subespècie "swordtail" no té
el locus lligat al X ni el locus del supresor autosòmic. Els descendents
dels retroencreuaments o els híbrids de la F2 poden heretar l'oncogen
sense el supresor.
L'estima de 9 o més loci responsables de
l'inviabilitat dels híbrids entre D. melanogaster i D.
simulans, les quals s'han separat fa molt de temps, pot ser una estima
per excés del nombre de loci que inicialment produïen aquesta
inviabilitat.
Podem concloure aquest aspecte dient que l'aïllament
reproductor entre espècies a vegades pot tenir una base genètica
senzilla.
EFECTE DEL CROMOSOMA X I LA REGLA DE HALDANE
Els cromosomes sexuals tenen un efecte molt important
sobre l'esterilitat i la inviabilitat dels híbrids.
Regla de Haldane: quan a la F1 de l'encreuament
entre dues espècies diferents un sexe no apareix, és rar
o estèril, aquest sexe és l'heterogamètic (Haldane,
1922).
S'ha suggerit que aquesta regla només reflecteix
el fet que els mascles són més sensibles que les femelles
a la hibridació, pero això no és cert doncs es compleix
tant en aquells taxons en que els mascles són el sexe heterogamètic
(mamífers, dípters) com en aquells en que les femelles són
el sexe heterogamètic (aus, lepidòpters). La regla té
molt poques excepcions.
A la taula següent es pot observar que la
major part dels casos d'hibridació entre espècies verifiquen
la regla de Haldane:
| Grup |
Caràcter |
Hibridacions amb asimetria
|
Verifiquen la regla de Haldane
|
| Mamífers |
Fertilitat |
20
|
19
|
| Aus |
Fertilitat |
43
|
40
|
| |
Viabilitat |
18
|
18
|
| Drosophila |
Fertilitat i viabilitat |
145
|
141
|
Així doncs, l'explicació de la regla
de Haldane ha d'estar relacionada amb els cromosomes sexuals i no amb el
sexe del'individu. Els gens que tenen un efecte més important sobre
la inviabilitat i la esterilitat dels híbrids són els gens
lligats al sexe.
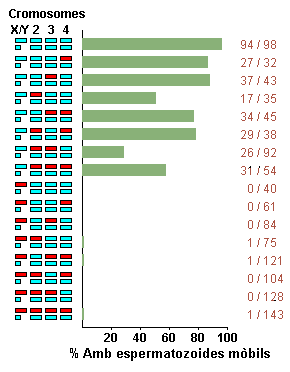 Un
experiment clàssic, que demostra la importància dels heterocromosomes
en la determinació de l'aïllament reproductor postzigòtic
és el realitzat per Orr (1987) amb les espècies Drosophila
pseudoobscura i D. persimilis. La hibridació entre aquestes
dues espècies produeix femelles fèrtils però mascles
estèrils. Quan es retrocreuen les femelles híbrides fertils
amb mascles de D. pseudoobscura s'obtenen alguns mascles
descendents fèrtils i d'altres estèrils. Utilitzant una soca
de D. pseudoobscura homozigòtica respecte a mutants
morfològics de cadscun dels 4 cromosomes, Orr va demostrar que tots
els cromosomes tenien un efecte significatiu sobre la fertilitat dels mascles
(mesurada segons la motilitat dels espermatozoides). Ara bé, l'efecte
més important el tenie el cromosoma X. Els mascles descendents del
retroentrecreuament, portadorsdel cromosoma X de
D. persimilis,
no tenien espermatozoids mòvils, eren doncs estèrils (figura
7).
Un
experiment clàssic, que demostra la importància dels heterocromosomes
en la determinació de l'aïllament reproductor postzigòtic
és el realitzat per Orr (1987) amb les espècies Drosophila
pseudoobscura i D. persimilis. La hibridació entre aquestes
dues espècies produeix femelles fèrtils però mascles
estèrils. Quan es retrocreuen les femelles híbrides fertils
amb mascles de D. pseudoobscura s'obtenen alguns mascles
descendents fèrtils i d'altres estèrils. Utilitzant una soca
de D. pseudoobscura homozigòtica respecte a mutants
morfològics de cadscun dels 4 cromosomes, Orr va demostrar que tots
els cromosomes tenien un efecte significatiu sobre la fertilitat dels mascles
(mesurada segons la motilitat dels espermatozoides). Ara bé, l'efecte
més important el tenie el cromosoma X. Els mascles descendents del
retroentrecreuament, portadorsdel cromosoma X de
D. persimilis,
no tenien espermatozoids mòvils, eren doncs estèrils (figura
7).
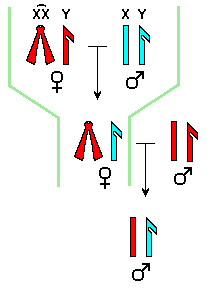 S'han
proposat dues explicacions per la regla de Haldane: Haldane, Dobzhansky
i Muller van proposar que els híbrids heterogamètics tenen
un desequilibri entre cromosomes X i dotacions d'autosomes: manca un cromosoma
X compatible amb una dotació d'autosomes. Perque l'híbrid
sigui viable o fertil cal que disposi d'un genoma haploide complet de cada
espècie. Una manera de comprovar si aquesta explicació és
correcta consisteix en obtenir femelles híbrides portadores dels
dos cromosomes X d'una espècie. Aquestes femelles hauríen
de ser també inviables o estèrils. Coyne i Orr van demostrar
que en algunes espècies de Drosophila que verificaven la
regla de Haldane, les femelles híbrides amb el mateix desequilibri
entre cromosomes X i autosomes que els mascles estèrils de la F1,
eren completament fèrtils.
S'han
proposat dues explicacions per la regla de Haldane: Haldane, Dobzhansky
i Muller van proposar que els híbrids heterogamètics tenen
un desequilibri entre cromosomes X i dotacions d'autosomes: manca un cromosoma
X compatible amb una dotació d'autosomes. Perque l'híbrid
sigui viable o fertil cal que disposi d'un genoma haploide complet de cada
espècie. Una manera de comprovar si aquesta explicació és
correcta consisteix en obtenir femelles híbrides portadores dels
dos cromosomes X d'una espècie. Aquestes femelles hauríen
de ser també inviables o estèrils. Coyne i Orr van demostrar
que en algunes espècies de Drosophila que verificaven la
regla de Haldane, les femelles híbrides amb el mateix desequilibri
entre cromosomes X i autosomes que els mascles estèrils de la F1,
eren completament fèrtils. 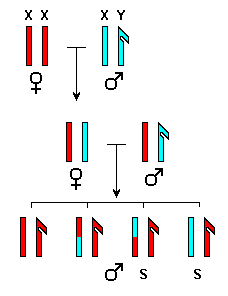 Per
tant, en el cas de la fertilitat, aquesta explicació de la regla
de Haldane no és correcta. Tanmateix, si que ho pot ser en el cas
de la inviabilitat dels híbrids: l'encreuament entre femelles de
D.
simulans i mascles de D. teissieri verifica la regla de Haldane:
s'obtenen només femelles híbrides. Els mascles híbrids
moren durant el primer o segón estadi larvari. Utilitzant una soca
de D. simulans amb els cromosomes X units, Orr (1993) ha demostrat
que les femelles híbrides desequilibrades (amb els dos cromosomes
X de D. simulans) són també inviables (figure 8).
Aquests resultats demostren que la base genètica de la regla de
Haldane és diferent en el cas de l'esterilitat dels híbrids
que en el cas de la inviabilitat dels híbrids. Un altre explicació
és que hi ha una incompabilitat entre loci del cromosoma X i loci
del cromosoma Y de les dues espècies progenitores.
Per
tant, en el cas de la fertilitat, aquesta explicació de la regla
de Haldane no és correcta. Tanmateix, si que ho pot ser en el cas
de la inviabilitat dels híbrids: l'encreuament entre femelles de
D.
simulans i mascles de D. teissieri verifica la regla de Haldane:
s'obtenen només femelles híbrides. Els mascles híbrids
moren durant el primer o segón estadi larvari. Utilitzant una soca
de D. simulans amb els cromosomes X units, Orr (1993) ha demostrat
que les femelles híbrides desequilibrades (amb els dos cromosomes
X de D. simulans) són també inviables (figure 8).
Aquests resultats demostren que la base genètica de la regla de
Haldane és diferent en el cas de l'esterilitat dels híbrids
que en el cas de la inviabilitat dels híbrids. Un altre explicació
és que hi ha una incompabilitat entre loci del cromosoma X i loci
del cromosoma Y de les dues espècies progenitores.
A les plantes l'al.loploidia és una causa
important d'especiació. També s'ha suggerit que l'aïllament
postzigòtic podria estar determinat perel moviment incontrolat d'elements
genètics transponibles en els híbrids, que normalment no
són mòvils dintre de l'espècie. Aquest model es basa
en el fenòmen de la disgènesi híbrida de D. melanogaster.
Un altre fenòmen interessant és el de la inviabilitat dels
híbrids causada per infeccions bacterianes: s'ha descrit en 5 ordres
d'insectes, es va descobrir al recuperar els híbrids inviables en
un medi amb antibiòtics (tots els casos inclouen el génere
de bacteris Wolbachia). La infecció d'una sola espècie
produeix la inviabilitat dels híbrids només en una direcció
de l'encreuament (efecte matern). Aquesta "especiació" per infecció
és instantània (com l'al.lopoliploidia). L'endosimbiosi s'ha
considerat per alguns autors com una causa general d'especiació
(simbiogènesi).
BIBLIOGRAFIA
COYNE, J. A. 1992. Genetics and Speciation. Nature,
355:511-515.
COINE, J. A. and ORR, H. A. 1989. Two rules of
speciation. In Speciation and its Consequences (eds Otte, D. & Endler,
J.): 180-207. Sinauer Associates Sunderland, Massachusetts.
DOBZHANSKY, TH. 1937. Genetics and the origin
of species. Columbia University Press. New York.
DOBZHANSKY, TH. 1970. Genetics of the Evolutionary
Process. Columbia University Press. New York.
FEDER, J. L., CHILCOTE, C. A. and BUSH, G. L.
1988. Genetic differentation betwen sympatric host races of the apple maggot
fly Rhagoletis pomonella. Nature, 336: 61-64.
GOULD, S. J. & Eldredge, N. 1977. Punctuated
equilibria: The tempo and mode of evolution reconsidered. Paleobiology,
3: 115-151.
HALDANE, J. B. S. 1922. Sex-ratio and unisexual
sterility in hibrids animals. J. Genet. 121:101-109.
KIMURA, M. 1983. The neutral theory of molecular
evolution. Cambridge University Press.
ORR, H. A. 1987. Genetics of male and female sterility
in hybrids of Drosophila pseudoobscura and D. persimilis.
Genetics, 116: 555-563.
ORR, H. A. 1993. Haldane's rule has multiple genetic
causes. Nature, 361:532-533.
PREVOSTI, A. 1993. Concepto de Especie en el Darwinismo
actual. Mundo Científico, 141:1040-1049.
RIDLEY, M. 1993. Evolution. Blackwell Scientific
Publications.