EVOLUCIÓ
Lluis Serra i Camo - curs 1998-1999
La mida del genoma seria constant si els únics mètodes
per produir canvis en el genoma fossin: deriva genètica, mutacions,
migracions. La grandària del genoma ha anat augmentant en el curs
de l'evolució així com la complexitat dels mecanismes que
actuen. En l'actualitat alguns mecanismes són ben coneguts; altres
no.
El "molecular drive" (deriva molecular) segons Dover engloba mecanismes
que provoquen mutacions no equivalents a les puntuals, de taxa elevada
i que provoquen canvis puntuals. Entre ells:
-
Transposició gènica
-
Entrecreuament desigual
-
Conversió gènica
-
Duplicació gènica
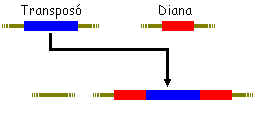
Transposició gènica
Existeixen seqüències de DNA que poden saltar dins del genoma
de una posició a una altre, són els elements mòbils
o elements transponibles. (De manera més estricta, gens saltadors
o transpossons).
Transmissió gènica horitzontal
Els bacteris poden adquirir DNA del medi i incorporar-lo al seu genoma
(transformació), es a dir, es produeix intercanvi entre espècies
diferents. També es pot produir entre procarionts i eucarionts o
entre eucarionts.
Les taxes són de 10^-2 a 10^-4 fenòmens per generació.
Existeixen dos tipus:
No replicativa - L'element transponible salta a una
altra posició denominada diana (la seqüència que hi
ha en aquesta regió queda duplicada abans i després del transpossó
????). En el lloc d'origen no queda la informació. Exemple els gens
saltadors del blat de moro descrits per la Barbara McKlintock.
Replicativa - Es produeix una duplicació del transpossó
i després un salt en el DNA. La diana també queda duplicada.
 |
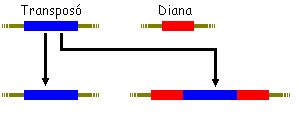 |
| Transmissió gènica no
replicativa
El fragment genètic blau desapareix del seu
lloc dorígen i es trasllada a una altre lloc del genoma on sinsereix. |
Transmissió gènica replicativa
El fragment genètic blau no desapareix
del seu lloc dorígen i quan es trasllada a una altre lloc del genoma
on sinsereix. |
Retrotranspossició
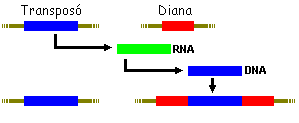
El transpossó prèviament es transcrit en RNA. ???? i
a apartir d'aquest es fa una copia en DNA que és el que es transpossa.
En aquest cas també queda còpia en el lloc original.
Alguns transpossons només són transpossats en casos concrets,
per exemple: el elements P de Drosophila son mòbils només
en cèl·lules germinals. Altres vegades s'insereixen sempre
en un mateix lloc del genoma, per exemple: L'operó de la galactosa
de E. coli. El fago Mu quan salta es pot transposar a qualsevol lloc del
genoma del bacteri. Els elements T? de Drosophila sempre salten al cromosoma
Y.
La retrotranspossició usa com intermediari RNA. En aquest cas
el promotor es transcriu gràcies a l'enzim RNA-polimerasa III. La
retrotranscriptasa inversa sintetitza el DNA a partir de l'RNA.
Etapes:
1 - Transcripció a RNA
2 - Retrotranscripció a híbrid DNA/RNA
3 - Copia del segon bri de DNA i degradació del RNA (per efecte
de la retrotranscriptasa inversa que també actua com RNA-asa)
4 - Transposició, inserció en el genoma.
Hi han molts tipus d'elements transponibles, els més complexos són
els retrovirus. Tots tenen tres gens com a mínim:
-
GAG - codifica una poliproteïna modificada postraduccionalment per
donar diverses proteïnes que formaren la capsida i la matriu ??? del
virió.
-
POL - codifica una poliproteïna que postraduccionalment es tallada
en tres fragments: una transcriptasa inversa, una integrasa (que permet
integrar el genoma viral en el de la cèl·lula) i una proteasa
que catalitza tots el processos de maduració de les poliproteïnes.
Per poder començar el procés, el virus porta dins seu
una molècula de proteasa procedent de la infecció anterior.
-
ENV - codifica glicoproteïnes de la coberta que reconeixen les cèl·lules
diana.
En diferents virus poden existir a més d'aquests altres gens; el
VIH per exemple, té 6 més.
Altres tipus d'elements transponibles són:
Retroelements són tots els que codifiquen la transcriptasa inversa;
no tots són transponibles. Així, els pararetrovirus no es
poden transposar, mentre que els retrovirus si.
Els retrotranspossons no tenen el gen ENV, pel que no poden formar proteïnes
de coberta i per tant no poden infectar altres cèl·lules
com a virions; però tot i així, són transponibles.
Els elements transponibles tenen importància evolutiva, ja que
al poder saltar donen lloc a canvis en el genoma, el que origina variabilitat
genètica.
Per exemple, el virus VIH té una taxa de mutacions de l'ordre
d'un milió de vegades més alta que la dels humans. Es degut
a que la transcriptasa inversa no té correcció d'errors (com
tenen les DNA-polimerases). Una taxa de mutació tan alta representa
un avantatge evolutiu, doncs li permet escapar als mecanismes de defensa
del cos humà.
 Aquesta
taxa tan alta es pot usar per seguir el procés de la infecció.
Per exemple, en el cas del dentista de Florida.
Aquesta
taxa tan alta es pot usar per seguir el procés de la infecció.
Per exemple, en el cas del dentista de Florida.
En 1990, al centre de control d'Atlanta arribà una noia que s'havia
infectat. Es va arribar a la conclusió de que l'infector era el
seu dentista, portador de la sida. Es va estudiar un tros del gen ENV de
virus obtinguts de 7 pacients suposadament infectats pel mateix dentista.
Amb les dades de la seqüenciació es va fer un arbre filogenètic.
Del dentista es van obtenir dues mostres, que degut a la pròpia
variabilitat del virus, eren diferents. Dels 7 pacients, 5 presentaven
seqüències molt properes a les del dentista. Hi havia 2 diferents,
en els que fallava la hipòtesis d'infecció pel mateix; eren
els únics dos que tenien risc d'haver-se infectat per altres medis:
l'un era drogaaddicte i l'altre homosexual.
Veiem que podem parlar d'evolució a una escala molt petita.
Entrecreuament desigual
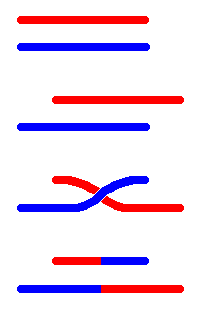 És
el principal mecanisme que explica la formació de nous gens (creixement
del genoma). Si podem duplicar gens, aquests poden adquirir noves funcions.
És
el principal mecanisme que explica la formació de nous gens (creixement
del genoma). Si podem duplicar gens, aquests poden adquirir noves funcions.
Per fer l'entrecreuament, generalment es produeix un aparellament entre
cromosomes (cromàtides homòlogues). De vegades, quan existeixen
seqüències semblants en llocs no homòlegs, les cromàtides
no s'aparellen bé, i es dóna l'entrecreuament desigual.
El resultat és que hi ha gens que es dupliquen en una cromàtida
i falten en l'altre, pel que l'una és més gran que l'altra.
Les duplicacions no solen ser perjudicials, però les pèrdues
són deletèrees.
Exemple: existeixen diversos tipus de globines humanes (lepore i antilepore,
el duplicat) generades per aquest mecanisme. Es produeix en mig de les
cadenes alfa i beta. Provat per que es detecten polipèptids híbrids.
Quan una part del genoma es duplica, poden passar diferents coses:
Que no es dupliqui la part de transcripció: el resultat és
un pseudogen, ja que no es pot transcriure. N'hi han molts.
Si conserva igual funció que l'original, el resultat pot ser
un increment en la quantitat de l'RNA i de la proteïna.
Un fragment duplicat pot experimentar mutacions que no l'inactivin,
assolint una nova funció, que pot ser o no propera a la del gen
original.
L'entrecreuament desigual es pot donar entre cromàtides germanes,
cromàtides de cromosomes homòlegs, i fins i tot, entre cromàtides
de diferents cromosomes.
Pot passar successives vegades. pel que resulten n còpies del
mateix fragment.
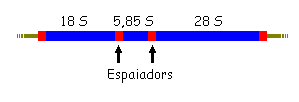 Exemple
1: gens de l'RNA ribosòmic en eucariotes. Un gen té la informació
del 18S, 5,8S i 28S seguides, donant un sol transcrit primari. En diverses
espècies hi ha diferent nº de còpies, i totes mantenen
la funció. Ex. en mitocondris humans, 1 còpia; en micoplasma,
2; en E. coli, 7; en Drosophila, 130-250; en Homo, ~300; i en Xenopus (gripau
africà), 400-600. És lògic per que calen molts ribosomes
especialment quan les cèl·lules es divideixen durant el creixement
embrionari.
Exemple
1: gens de l'RNA ribosòmic en eucariotes. Un gen té la informació
del 18S, 5,8S i 28S seguides, donant un sol transcrit primari. En diverses
espècies hi ha diferent nº de còpies, i totes mantenen
la funció. Ex. en mitocondris humans, 1 còpia; en micoplasma,
2; en E. coli, 7; en Drosophila, 130-250; en Homo, ~300; i en Xenopus (gripau
africà), 400-600. És lògic per que calen molts ribosomes
especialment quan les cèl·lules es divideixen durant el creixement
embrionari.
Exemple 2: cas de les histones, responsables del enrrotllament de l'DNA
i de l'estructuració en solenoide. S'han conservat al llarg de l'evolució
i amb la mateixa funció per a tots els organismes (de la més
nova a la més vella, només hi ha un aa de diferència).
Estan en tàndem en el genoma i hi ha diferent nº de còpies.
Ex: garota, de 300-3000 còpies; Xenopus, 20-50; Homo, 30-40.
Conversió gènica
Té com a conseqüència evolutiva la homogeneització
de seqüències nucleotídiques. Es produeix quan es reparen
els productes de recombinació.
El model de Holiday de recombinació a nivell molecular és
el més acceptat actualment (és antic).
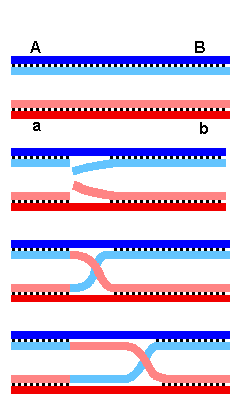
Dues cadenes de DNA pertanyents a dues cromàtides homòlogues
es col·loquen enfrontades i de forma que els brins en contacte tinguin
la mateixa polaritat.
Una ribonucleasa????? talla els dos brins que estan en contacte, a la
mateixa alçada.
Una ligasa enganxa cada bri amb l'extrem de l'altre, formant una estructura
creuada en forma de ji, amb una cadena superposant-se a l'altre.
Si el creuament (ji) es desplaça lateralment, es formen regions
heteroduplex de parelles de bases no complementàries, per tenir
origen diferent.
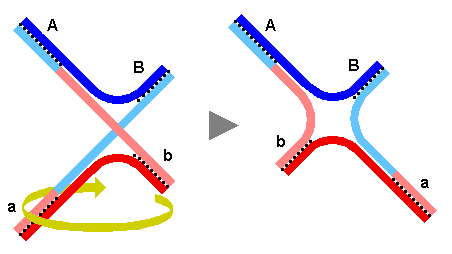
Tota l'estructura s'obre per rotació de 180º de una meitat.
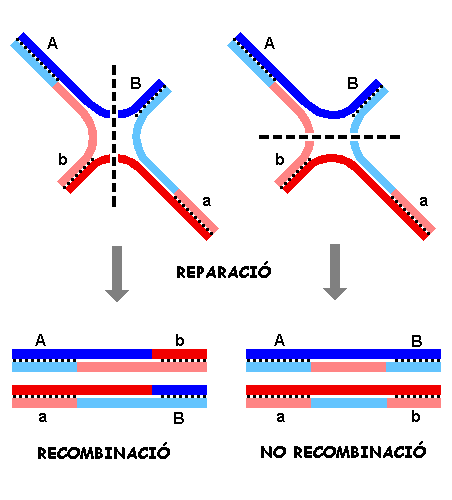
Es poden produir talls en direcció N-S o E-W. Un cop efectuada
la unió de les cadenes tallades podem considerar dos possibilitats.
Cas N-S: Hi ha recombinació i fragment heteroduplex.
Cas E-W: Resulta una estructura no recombinant però que conté
un fragment heteroduplex.
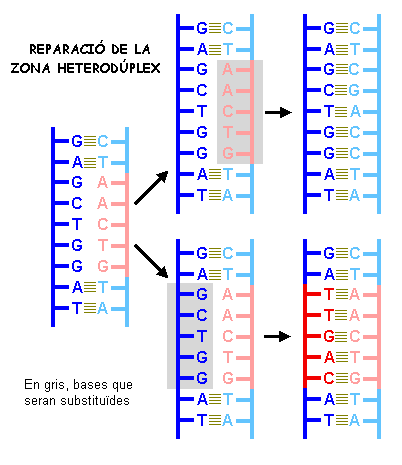
A nivell molecular el crossing-over porta a la aparició de regions
heteroduplex, que poden ser molt grans, d'una longitud de mil·lers
de parells de bases i que afecten com a mínim a un gen. La cèl·lula
repara aquestes regions per fer-les complementàries.
El mecanisme de reparació consisteix en substituir les bases
d'una cadena per les complementàries de l'altra. Segons quina cadena
es prengui com motlle poden produir-se canvis genètics.
Cas de les asques de Neurospora
Neuropora és un fong amb hifes haploids i cicle biològic
haplont. Les espores es formen després d'una meiosi seguida d'una
mitosi (en cada asca es formen 8 espores). Espècie usada per estudis
de meiosi creuant filaments genèticament diferents. Un d'aquests
caràcters és la pigmentació de les espores, que poden
ser blanques o negres. El resultat més freqüent és 4
d'un color seguides de 4 de l'altre.  Sovint apareixen ordenacions diferents,
Sovint apareixen ordenacions diferents,  però mantenint les proporcions; es donen quan ha hagut recombinació.
Molt rarament, apareixen asques amb proporcions diferents: 5 i 3, 6 i 2
o 7 i 1,
però mantenint les proporcions; es donen quan ha hagut recombinació.
Molt rarament, apareixen asques amb proporcions diferents: 5 i 3, 6 i 2
o 7 i 1, 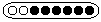 aparentment inexplicables
i d'heterodúplex.
aparentment inexplicables
i d'heterodúplex.
Si els heterodúplex no han estat corregits al produir-se la mitosi,
ja no es corregeixen, doncs cada cadena forma la seva complementària
en la replicació, i desapareix l'heterodúplex. En aquest
cas el resultat és el normal.
De vegades, entre la meiosi i la mitosi final es corregeixen els heterodúplex
d'alguns nuclis o de tots ells; en aquests casos, si els fragments heterodúplex
afecten al gen que determina el color de les espores, es poden obtenir
les combinacions atípiques. Aquestes apareixen al obtenir-se més
nuclis amb la informació de la cadena usada de motlle en la reparació
que nuclis amb la informació que s'ha descartat. Així, si
només s'ha reparat l'heterodúplex d'un dels nuclis, obtindrem
5 i 3. Si es reparen dos i en tots dos s'ha descartat la mateixa cadena,
6 i 2, i així successivament.
La reconversió generalment és esbiaixada. Uns al·lels
tenen més tendència que altres a ser reparats, això
porta a homogeneitzar seqüències. El al·lels que es
reparen més tenen tendència a desaparèixer.
Existeix conversió al.lèlica dins de la mateixa cromàtida,
entre cromàtides diferents/germanes però no al.leliques i
fins i tot entre gens de cromosomes no homòlegs (ectòpica).
En resum, la conversió gènica:
-
S'ha observat en tots els loci.
-
Es presenta amb una freqüència del 0,5 - 18 % (varia segons
loci). És molt alta.
-
No sempre es reparen els al.lels.
-
El biaix fa que els dos al·lels no siguin equiprobables.
-
La mida de la regió heteroduplex pot oscil·lar entre uns
pocs parells de bases i més de 12.000.
Duplicació gènica
Nivells a que es dóna:
Una part d'un gen (gènica parcial o interna).
Gens complets (gènica completa)
Part d'un cromosoma (cromosòmica parcial)
Tot un cromosoma (aneuploidia)
Tot el genoma (poliploidia) Això ha estat freqüent en animals
fins l'aparició dels mecanismes cromosòmics de determinació
genètica del sexe, la selecció natural elimina.... En animals
és relativament poc important. Al món vegetal, que no té
determinació sexual cromosòmica (moltes espècies són
hermafrodites), l'alopoliploïdia ha estat un fenomen molt freqüent.
Duplicació interna o parcial de gens
Un domini d'una proteïna es una regió d'aminoàcids
amb estructura ben definides (per exemple en proteïnes transmembranals,
formen les parets dels canal iònics). Els dominis funcionals tenen
una funció determinada, com unir-se a un substrat; no hi ha proximitat
entre els aa que el constitueixen, que poden trobar-se escampats per tota
la proteïna.
En proteïnes globulars existeix correspondència biunívoca
entre les posicions dels exons del gen i els dominis de les proteïnes.
Casos:
-
1 exó codifica exactament un domini.
-
1 exó codifica aproximadament un domini.
-
1 exó codifica més d'un domini.
-
Mes de 1 exó codifica un domini.
-
Cap relació
Les duplicacions gèniques internes poden donar-se per entrecreuaments
asimètrics entre introns que presentin similituds.
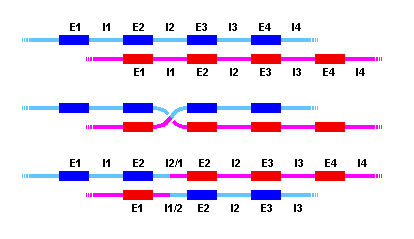 E1--I1--E2--I2--E3--I3--E4--I4--
E1--I1--E2--I2--E3--I3--E4--I4--
E1--I1--E2--I2--E3--I3--
Si I1 i I2 tenen semblança, es pot donar entrecreuament entre
ells, resultant:
E1--I1--E2--I2/1--E2--I2--E3--I3--E4--I4--
E1--I1/2--E3--I3--
El resultat és que un exon s'ha duplicat en una de les cromàtides
i perdut en l'altra (sol ser per aquesta segona una mutació deletèrea).
L'increment d'un exon pot fàcilment ser funcional, i de vegades
incrementar la seva eficàcia.
Hi ha exemples que demostren que és un mecanisme molt freqüent,
i més en vertebrats que en invertebrats.
Exemple: L'Haptoglobina humana (proteïna encarregada de reciclar
el Fe en la degradació de l'Hb). Es tracta d'una seqüència
de 84 aminoàcids. Existeixen dos al.lels: a1(F)
i a1(S), separables per electroforesi (F = fast,
S = slow). Existeix un altre al.lel a2 que codifica
una mol.lècula de 143 aminoàcids, amb una seqüència
híbrida de a1(F) i a1(S).
Es tractaria d'una modificació posterior a la separació evolutiva
de la branca humana de la dels ximpanzés (fa entre 4 i 5 milions
d'anys) doncs aquests animals no la posseeixen. L'al.lel
a2 es presenta amb una freqüència des 30 - 70 % en els
caucàsics i pot acabar estenent-se per la seva major eficàcia,
ja que presenta més afinitat pel Fe que les formes a1
i es tracta doncs d'una modificació beneficiosa.
Exemple: El col.lagen està format per tres cadenes polipeptídiques
enrotllades helicoidalment, amb una repetitivitat d'aa molt alta.
El gen a2 del col·lagen de tipus I
del pollastre té 38 Kbases i 52 exons (molts!!!) la part del gen
que codifica l'hèlix de col.lagen té 42 exons.
5 de 45 parells de bases
23 de 54 parells de bases
5 de 99 parells de bases
8 de 108 parells de bases
1 de 162 parells de bases
Cada hèlix té 338 tríades de aminoàcids del
tipus Gly-X-Y (preferentment X = prolina i Y = hidroxiprolina). Cada aminoàcid
ve determinat per tres bases, una tríada per 9. En total, cada hèlix
es codifica per 338 * 9 =3042 bases.
Tots els grups 45, 54, 99... són divisibles per 9.
L'exó de 54 pb està molt repetit. Podria ser l'exó
ancestral que per entrecreuament desigual produït diverses vegades
ha donat el gen resultant.
El gen del col.lagen s'ha produït per repetició d'un gen
ancestral de 54 parells de bases.