|
Procesos anabólicos
y catabólicos
Sin duda, las células son
los laboratorios más sofisticados que existen. En un espacio increíblemente
reducido tienen lugar multitud de reacciones químicas, todas ellas
perfectamente coordinadas y reguladas. Muchas de esas reacciones son además
mutuamente incompatibles, es decir, si trituramos la célula y se mezclan
sus componentes el resultado es un caos metabólico. Para que las
diferentes rutas metabólicas operen en armonía, es imprescindible un
control riguroso mediante diferentes enzimas
específicas, y que las distintas rutas ocurran, en muchos casos, en
compartimentos celulares separados (orgánulos celulares).
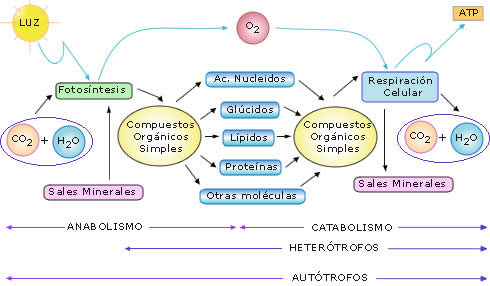
Todos los procesos metabólicos se pueden clasificar en dos tipos:
procesos anabólicos,
o de síntesis, y procesos
catabólicos, o de degradación.
Se puede decir que el anabolismo se inicia con la síntesis de los
primeros compuestos orgánicos a partir de sustancias inorgánicas,
mediante la fotosíntesis o la quimiosíntesis.
Esos primeros pasos anabólicos sólo los pueden realizar los organismos
autótrofos. Luego, a partir de moléculas orgánicas simples, se formarán,
mediante diferentes rutas anabólicas, todos los componentes orgánicos de
los seres vivos.
El catabolismo se puede iniciar con la descomposición de muy diferentes
sustancias orgánicas, pero, al final, la mayoría de las rutas catabólicas
confluyen en la respiración celular, a través de la cual los compuestos
orgánicos se terminan por degradar en sustancias inorgánicas.
Naturalmente, muchas de las reacciones químicas, tanto anabólicas como
catabólicas, implican transformaciones energéticas, y los procesos que
liberan energía (en general los catabólicos) se acoplan a los que la
consumen (en general los anabólicos)
|
|
|
Figura 1: Esquema simplificado de los principales procesos anabólicos
y catabólicos realizados por los organismos autótrofos y heterótrofos
|
2.
La fotosíntesis
2.1. Modalidades y fases de la fotosíntesis
La fotosíntesis es el proceso mediante el cual las sustancias inorgánicas
simples (CO2 , H2O y, por extensión, nitratos y
sulfatos) se combinan para formar compuestos orgánicos simples,
utilizando para ello la energía de la luz.
Se pueden diferenciar dos modalidades de fotosíntesis:
-
Fotosíntesis
oxigénica.
Se denomina así porque en ella se desprende O2 (a partir del H2O
). Es la que realizan las plantas, las algas y las cianobacterias.
-
Fotosíntesis anoxigénica.
Llamada así porque en ella no se libera O2, ya que el agua no
interviene como dadora de electrones. Existen diferentes modalidades y la
realizan algunas bacterias sulfúreas y no sulfúreas.
En adelante nos centraremos únicamente en el estudio de la fotosíntesis
oxigénica.
El conjunto de procesos que tienen lugar en la fotosíntesis vegetal se
puede resumir en la siguiente ecuación:
energía luminosa
6 CO2 + 6 H2O ----------------------> C6H12O6
+ 6 CO2
clorofila
Pero
el agua no puede reaccionar directamente con el CO2, luego
entre el sustrato inicial y los productos finales de esa reacción deben
ocurrir complejos procesos metabólicos. Esos procesos se dividen en dos
fases:
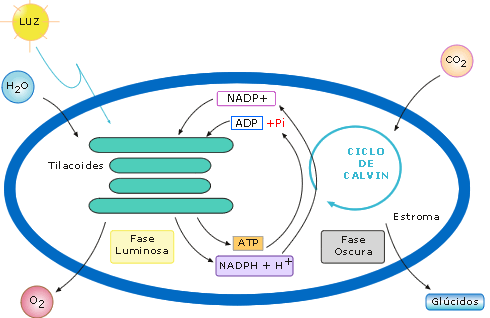
-
Fase luminosa. Ocurre en la membrana tilacoidal de los cloroplastos. En
ella la energía de la luz impulsa la formación de poder energético, en
forma de ATP, y poder reductor, en forma de NADPH.
- Fase oscura. Ocurre en el estroma de los cloroplastos. En ella la
energía del ATP y el NADPH, obtenidos anteriormente, impulsan la
reacciones para la formación de compuestos orgánicos simples a partir de
sustancias inorgánicas.
|
|
|
Figura 2: Esquema de la fase luminosa y la fase oscura de la fotosíntesis
|
2.2.
Obtención de energía: fase luminosa
¿Cómo
es captada la energía luminosa?
La energía de la luz es captada por los pigmentos fotosintéticos
situados en la membrana tilacoidal de los cloroplastos.
Al incidir un fotón
sobre un pigmento fotosintético, desplaza un electrón hacia un nivel de
mayor energía. El pigmento excitado puede volver a su estado original de
tres formas:
-
Perdiendo la energía extra en forma de luz y calor (fluorescencia).
- Mediante una transferencia de energía por resonancia, en la que la
energía (pero no el electrón) pasa de un pigmento a otro.
- Mediante una oxidación
del pigmento, al perder el electrón de alta energía, que será captado
por un transportador de electrones.
El
pigmento fotosintético más importante es la clorofila, que absorbe el
color violeta, azul y rojo, y refleja el verde. Además existen otros
pigmentos accesorios, como los carotenoides, que reflejan el rojo,
anaranjado o amarillo.
|
Cuestión para autoevaluación:
|
|
¿Por qué en otoño, al degradarse la clorofila de las
hojas de los árboles caducifolios, éstos adquieren tonalidades
pardas, amarillentas y rojizas?
|
|
Orientación
|
|
Porque al desaparecer la clorofila
queda desenmascarado el color de los otros pigmentos carotenoides
|
Todos los pigmentos fotosintéticos se agrupan en fotosistemas
que, hipotéticamente, podemos imaginar con forma de embudo. Pueden ser de
dos tipos: fotosistema I (con un máximo de absorción de 700 nm) y
fotosistema II (con un máximo de absorción de 680 nm).

Figura 3: Esquema de un fotosistema
con su complejo antena
y centro de reacción fotoquímico
2.2.
Obtención de energía: fase luminosa
¿Cómo
fluyen los electrones de un transportador a otro?
Los
electrones que salen del centro de reacción fotoquímico pueden seguir
dos recorridos: flujo no cíclico y flujo cíclico de electrones.
-
Flujo no cíclico de electrones
En
él intervienen los dos fotosistemas, y se denomina así porque los
electrones que salen de las moléculas de clorofila ya no regresarán a
esas mismas moléculas. Aunque la secuencia de acontecimientos puede ser
diferente, simplificando, consideraremos que se inicia con la
fotólisis de agua: un proceso básico en el que una molécula
de agua se escinde en 2e-, 2H+ y ½ O2. Los electrones pasan la
fotosistema II, que previamente habrá perdido otros dos electrones por la
acción de dos fotones de luz. Los dos electrones del fotosistema II
alcanzan un nivel energético alto, y luego fluyen, a través de varios
transportadores, hacia el fotosistema I, que previamente habrá perdido
otros dos electrones por la incidencia de dos fotones. Los electrones
"energéticos" del fotosistema I son captados por el aceptor
de electrones ferredoxina, y luego el NADP+, que se convierte
en NADPH + H+ (poder reductor).
Al mismo tiempo que ocurren estos procesos, el flujo de electrones hacia
niveles energéticos más bajos, impulsa el acúmulo de protones en el
espacio tilacoidal, generando un gradiente electroquímico o fuerza
protón-motriz. Luego, los protones salen del espacio
tilacoidal "cuesta abajo", a través de las ATP
sintasas, y hacen posible la fotofosforilación,
con la consiguiente formación de ATP (poder energético) a partir de ADP
y Pi.
Como subproducto de todo este proceso queda libre un átomo de oxígeno.
|

|
|
Figura 4: Esquema del flujo no cíclico de electrones
|
.2.
Obtención de energía: fase luminosa
-
Flujo cíclico de electrones
En él interviene únicamente el fotosistema I y se denomina así porque
los electrones que salen de la clorofila del fotosistema regresan de nuevo
a las mismas moléculas.
La única finalidad de este flujo es formar un poco más de ATP,
necesario, como veremos, para compensar el déficit que se produciría en
caso contrario, durante la fase oscura.
Los electrones salen cargados de energía del fotosistema, pasan por
varios transportadores de electrones, e impulsan indirectamente la
fotofosforilación, con la consiguiente formación de ATP.

Figura 5: Esquema del flujo cíclico de electrones
|
Cuestión para autoevaluación:
|
|
¿Son muy diferentes los transportadores de electrones
del flujo cíclico y del flujo no cíclico?
|
|
Orientación
|
|
No, son los mismos transportadores;
lo que ocurre es que la ferredoxina los cede al complejo b6-f
en vez de pasarlos al NADP+
|
2.3. Síntesis de materia orgánica: fase oscura
Gracias a la energía
obtenida en forma de poder reductor (NADPH) y poder energético (ATP) en
la fase luminosa, ahora será posible la fijación del carbono inorgánico
(CO2) en carbono orgánico (glúcidos simples). Además, por
otras vías metabólicas, también se podrán fijar, en forma de
compuestos orgánicos, los nitratos (NO3-) y
sulfatos (SO42-) inorgánicos.
La fijación del carbono tiene lugar a través del ciclo de Calvin o ciclo
C3, ya que la mayoría de los metabolitos
intermediarios tienen tres carbonos. Básicamente, en este ciclo se pueden
diferenciar tres etapas:
- Carboxilación.
La enzima rubisco cataliza la combinación de la ribulosa difosfato con el
CO2, formándose un compuesto intermedio e inestable (de 6 átomos
de carbono), que se descompone en dos moléculas de fosfoglicerato (con 3
átomos).
- Reducción. Mediante la energía que suministra el ATP y el poder
reductor del NADPH, el fosfoglicerato se transforma en gliceraldehído
3-fosfato.
- Recuperación. De cada 6 moléculas de gliceraldehído 3-fosfato,
5 se transforman en 3 moléculas de ribulosa difosfato (con consumo de
ATP) y la otra se considera el rendimiento neto del ciclo.
En definitiva, por
cada tres vueltas del ciclo de Calvin, 3 moléculas de CO2 se
combinan al hidrógeno de 6 NADPH, impulsadas por la energía de 9 ATP,
obteniéndose como primer compuesto orgánico una molécula de gliceraldehído
3-fosfato.

Figura
6: Esquema del ciclo de Calvin. Por cada tres vueltas se forma como
rendimiento neto una molécula de gliceraldehído 3-fosfato.
2.4. Los productos de la fotosíntesis
La primera molécula
orgánica que se forma en la fotosíntesis, a partir del ciclo de Calvin,
es el gliceraldehído 3-fosfato. Luego, esta molécula será la precursora
de diferentes tipos de moléculas orgánicas, algunas de las cuales únicamente
tendrán C, H y O, mientras que otras tendrán además N o S orgánico.
- Para la síntesis
de compuestos orgánicos con carbono basta con el gliceraldehído.
Lo más común es que dos moléculas de gliceraldehído se unan formando
un molécula de glucosa, que se suele considerar como el producto final de
la fotosíntesis.

Figura
7: Rendimiento neto del proceso de fotosíntesis para obtener
una molécula de glucosa
-
Para la síntesis de compuestos orgánicos con nitrógeno, además del
gliceraldehído será necesario reducir los nitratos (NO3-),
para incorporarlos a la materia orgánica.
En este proceso se
diferencian tres etapas: a) reducción
de nitratos a nitritos, b) reducción de nitritos a amoníaco, y c)
incorporación del amoníaco al aminoácido glutamina. En las plantas este
proceso ocurre fundamentalmente en los cloroplastos y requiere energía
que es aportada por el ATP y el NADPH, obtenidos a partir de la fase
luminosa de la fotosíntesis.

Figura 8:
Para sintetizar compuestos orgánicos con nitrógeno, las plantas
deben reducir los nitratos inorgánicos
2.5. Factores que influyen en la actividad de la
fotosíntesis
El rendimiento de la
actividad fotosintética es influenciado por varios factores.
Los más importantes son:
- Intensidad
luminosa. En
general, a mayor intensidad luminosa, mayor actividad fotosintética.
Pero, cada especie está adaptada a unos niveles de iluminación óptima,
de intensidad variable. Si se superan esos niveles, se llega a la saturación
lumínica e, incluso, podrían deteriorarse los pigmentos fotosintéticos.
Por otro lado, también influye el color de la luz: el mejor es el que
absorbe (y no refleja) la clorofila.
- Temperatura. Como norma general, a mayor temperatura, mayor
actividad fotosintética, hasta que se llega a un máximo, superado el
cual se pueden desnaturalizar algunas enzimas. La temperatura óptima
variará de unas especies a otras.
- Concentración de CO2. A mayor concentración de CO2
mayor actividad fotosintética, hasta que se llega a un punto en el que se
estabiliza.
- Concentración de O2. Al aumentar la concentración de
O2 baja el rendimiento de la fotosíntesis debido a la fotorrespiración.

Figura
9: Influencia de diversos factores sobre la actividad fotosintética
3. Los procesos catabólicos
3.1. Esquema general del catabolismo
En el curso de los
procesos catabólicos las moléculas orgánicas se van degradando, paso a
paso, hasta formar otras moléculas más simples y, finalmente, sustancias
inorgánicas. Globalmente son procesos de oxidación
en los que las moléculas orgánicas van perdiendo electrones que, tras
pasar por una cadena transportadora, serán captados por una molécula
aceptora de electrones final. En esos procesos la energía liberada
permite la formación de moléculas de ATP. Según quien sea el aceptor
final de electrones se pueden diferenciar dos modalidades:
- Fermentación.
El aceptor final de electrones es un compuesto orgánico, por lo que se
trata de una oxidación incompleta y un proceso anaerobio.
- Respiración celular. El aceptor final de electrones es una
sustancia inorgánica. Si es el O2, se trata de una respiración
aerobia
que realizan la mayoría de los organismos; si es otro compuesto inorgánico
(NO3-, SO42-, CO2),
se trata de una respiración anaerobia, exclusiva de ciertos
microorganismos.
Los procesos catabólicos
se pueden iniciar a partir de varios sustratos orgánicos, especialmente
glúcidos, lípidos y proteínas. En cualquier caso, al final, las
diferentes rutas metabólicas de la respiración celular confluyen en el
ciclo de Krebs, y los electrones liberados en el proceso pasan a la cadena
respiratoria para formar ATP por fosforilación oxidativa. Una variante a
este esquema general es el desvío o la ruta alternativa de las
fermentaciones.
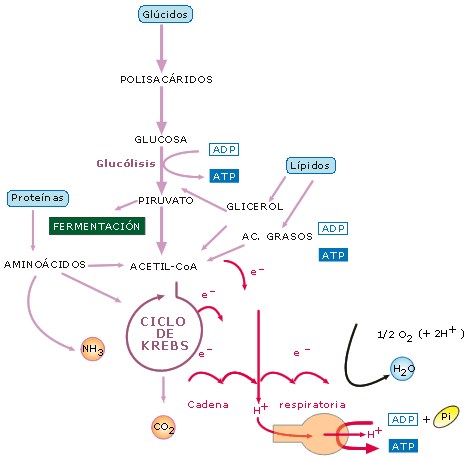
Figura 10: Esquema simplificado del catabolismo. Al tiempo que los
compuestos orgánicos se descomponen en CO2 y H2O,
los electrones son transferidos a lo largo de la cadena respiratoria
impulsando la fosforilación oxidativa
3.2. El ciclo de Krebs
Casi todos los
compuestos orgánicos (azúcares y grasas especialmente) se descomponen
hasta formar acetil-CoA: un grupo acetilo de dos carbonos, unidos a la
coenzima A.
El acetil-CoA ingresa ahora en una secuencia cíclica de reacciones químicas
que se conocen con el nombre de ciclo de Krebs o ciclo del ácido cítrico.
Este ciclo tiene lugar en la matriz mitocondrial y consiste en una
secuencia de ocho reacciones consecutivas. En el conjunto de esas
reacciones tienen lugar, básicamente, los siguientes acontecimientos:
- Se liberan 2 moléculas
de CO2, que se puede considerar que proceden del grupo acetilo
del acetil-CoA, con lo cual se consigue una oxidación completa la materia
orgánica. El CO2 será, por tanto, el producto residual último
de la respiración aerobia.
- Los electrones más los protones (hidrógeno), que pierde el grupo
acetilo al oxidarse, van a parar, en cuatro fases, a transportadores de
electrones, formándose 3 moléculas de NADH + H+ y una molécula
de FADH2. El poder reductor de estas moléculas será
transferido luego a través de la cadena respiratoria mitocondrial.
- En un paso del ciclo tiene lugar una fosforilación a nivel de sustrato
que origina una molécula de GTP (equivalente desde el punto de vista
energético a un ATP)
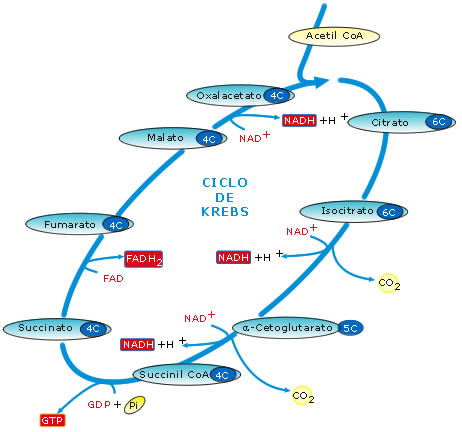
Figura
11: Esquema simplificado del ciclo de Krebs
3.3. La
cadena respiratoria
Los
transportadores de electrones NADH y FADH2, originados
fundamentalmente en el ciclo de Krebs, pero también en otros procesos
catabólicos, albergan el poder reductor que les confieren los electrones
"energéticos" que transportan.
Esa energía será liberada, poco a poco, a lo largo de la cadena
respiratoria que tiene lugar en las crestas y en la membrana mitocondrial
interna. En dicha membrana existen tres complejos enzimáticos
transportadores de electrones:
-
El complejo NADH deshidrogenasa
- El complejo citocromo b-c1
- El complejo citocromo oxidasa.
Tanto
el NADH como el FADH2 ceden los electrones "energéticos"
a la cadena formada por esos tres transportadores y, a medida que pasan de
un transportador a otro, los electrones van liberando energía. Esa energía
(según la teoría quimiosmótica de Mitchell) permite el bombeo de
protones desde la matriz mitocondrial al espacio intermembranoso de la
mitocondria. De este modo se genera un gradiente electroquímico de
protones, con una concentración de protones mayor en el espacio
intermembrana que en la matriz.
La fuerza
protón-motriz generada, impulsa los protones a través de las ATP
sintasas presentes en la membrana mitocondrial interna,
permitiendo la unión del ADP a un grupo fosfato, con la consiguiente
formación de ATP. El conjunto de estos procesos, que culminan con la
formación de ATP, constituyen la fosforilación
oxidativa.
Con
fines prácticos, aunque no es del todo exacto, se considera que una molécula
de NADH permite la formación de 3 moléculas de ATP, mientras que una de
FADH2 sólo aportará 2 ATP
Tanto los electrones como los protones, que han sido impulsados a lo largo
de la cadena respiratoria, deben unirse a un aceptor final. En la
respiración aerobia el aceptor
último de electrones (y protones) es el O2, que al
unirse al H2, forma H2O como producto final.

Figura 12: Transferencia de
electrones y fosforilación oxidativa
en la cadena respiratoria mitocondrial
|
Cuestión para autoevaluación
|
|
¿Cual es el destino último del O2 que
necesitamos inspirar de forma continua?
|
|
Orientación
|
|
|
3.4. El catabolismo de los glúcidos
Los glúcidos se consideran nutrientes energéticos (1 g
de glúcidos aporta como promedio 4 kcal). En su catabolismo, primero los
polisacáridos se descomponen hasta formar finalmente glucosa.
Concretamente, en los animales, el glucógeno acumulado en las células
hepáticas o en las fibras musculares se va hidrolizando por un proceso de
glucogenolisis.
La glucosa entra a continuación en una secuencia de
reacciones en las que se diferencian:
Glucólisis.
Consiste en 10 reacciones consecutivas, que tienen lugar en el citosol,
en las que se diferencian dos fases:
a. Fase preparatoria. Se consumen 2 moléculas de ATP para transformar una de
glucosa en 2 de gliceraldehído 3-fosfato.
b. Fase de beneficio. Se obtienen 4 moléculas de NADH y 4 moléculas de ATP,
formándose al final de la glucólisis 2 de piruvato.
En
el conjunto de la glucólisis, a partir de cada molécula de glucosa, se
obtienen, por tanto, como rendimiento neto 2 ATP (por fosforilación
a nivel de sustrato), y 2 NADH que luego podrán pasar a la
cadena respiratoria.
Figura 13: Esquema simplificado de la glucólisis
El
piruvato que se obtiene al final de la glucólisis se encuentra en una
encrucijada metabólica en la que puede seguir dos destinos, dependiendo
de la disponibilidad de oxígeno y el tipo de célula. Puede seguir la vía
anaerobia
de las fermentaciones
o la vía aerobia
de la respiración
celular.
-
Por la vía aerobia de la respiración celular el piruvato pasa al
interior de la mitocondria, donde, mediante una reacción irreversible, se
une a una coenzima
y sufre una descarboxilación (pérdida de CO2) y una oxidación,
formándose: CO2, NADH y acetil-CoA.
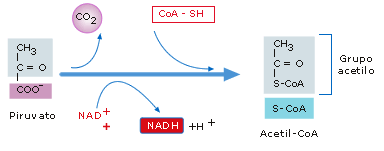
Figura 14: Transformación del piruvato en acetil-CoA
-
El NADH liberará su poder reductor en la cadena
respiratoria, mientras que el acetil-CoA ingresará en el
ciclo de Krebs.
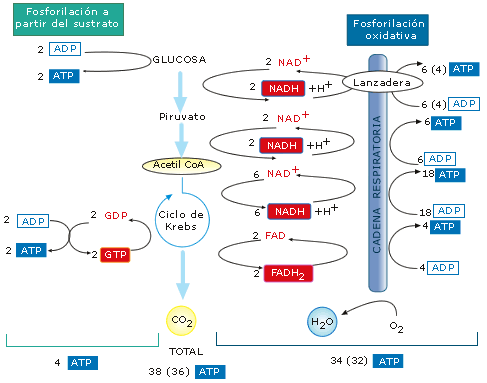
Figura 15: Balance energético
global de la respiración aerobia de
una molécula de glucosa
|
Cuestión para autoevaluación
|
|
¿En qué proceso metabólico, y mediante qué modalidad
de fosforilación, se obtiene el ATP que se forma en las
fermentaciones?
|
|
Orientación
|
|
|
3.5.
La vía anaerobia de las
fermentaciones
El
piruvato formado en la glucólisis no siempre sigue la vía de la
respiración celular; en determinadas circunstancias puede pasar a la vía
alternativa de las fermentaciones.
Las fermentaciones
genuinas son procesos anaerobios,
realizados por microorganismos que no toleran el oxígeno o por ciertas células
animales o vegetales cuando no disponen de suficiente oxígeno. Son poco
rentables desde el punto de vista energético, ya que la oxidación de la
materia orgánica es incompleta y se forma mucho menos ATP que en la
respiración celular aerobia. En general, únicamente 2 ATP por cada molécula
de glucosa.
Dependiendo el producto final, se diferencian varios tipos de
fermentaciones. Las más importantes son:
-
Fermentación alcohólica. En ella el piruvato se transforma en etanol y se
desprende CO2. La realizan, sobre todo, levaduras del género Saccharomyces
que tienen interés en la industria alimenticia por los productos
residuales de su metabolismo: el CO2 para esponjar la masa en
la fabricación del pan; y el etanol para producir diferentes bebidas
alcohólicas (vino, sidra, cerveza...).
- Fermentación láctica. En ella el piruvato se transforma en
lactato. La realizan diversas bacterias (Lactobacillus...) que
fermentan la leche, y se utilizan para obtener derivados lácteos. Por
otro lado, también la pueden llevar a cabo las células musculares cuando
no reciben suficiente oxígeno. Así, cuando se realiza un esfuerzo
intenso y prolongado, los músculos obtienen un poco de energía extra sin
necesidad de oxígeno, recurriendo a la fermentación; pero las
consecuencias de este proceso serán, posteriormente, las agujetas.
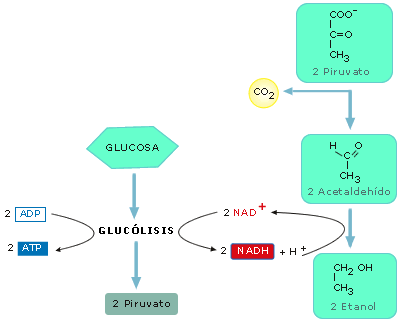
Figura 16-a: Fermentación alcohólica
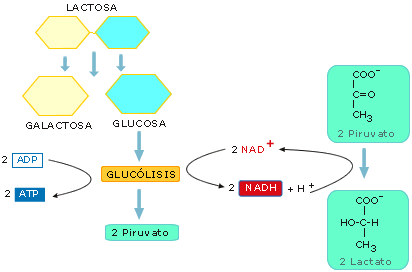
Figura 16-b: Esquema de la fermentación láctica
|
Cuestión para autoevaluación:
|
|
¿En qué proceso metabólico, y mediante qué modalidad
de fosforilación, se obtiene el ATP que se forma en las
fermentaciones?
|
|
Orientación
|
|
|
3.6. El catabolismo de los lípidos
Los lípidos, y más
concretamente las grasas o acilglicéridos, aportan aún más energía que
los glúcidos (1 g de grasas aportan 9 kcal).
En primer lugar, los acilglicéridos se descomponen en glicerol y en ácidos
grasos.
El glicerol puede seguir varios destinos metabólicos, pero en el catabólico
se transforma en dihidroxiacetona-fosfato y se incorpora a la glucólisis,
siguiendo las rutas posteriores ya conocidas. Al final, la respiración
aerobia de una molécula de glicerol puede proporcionar hasta 22 ATP.
Los ácidos
grasos pasan al interior de las mitocondrias y, en la matriz
mitocondrial, tiene lugar la ß-oxidación de los ác. grasos. En este
proceso cada ác. graso se va oxidando, liberándo, paso a paso,
"fragmentos" de dos carbonos hasta que termina por consumirse.
En cada paso se forman una molécula de FADH2, una de NADH y un
acetil-CoA.
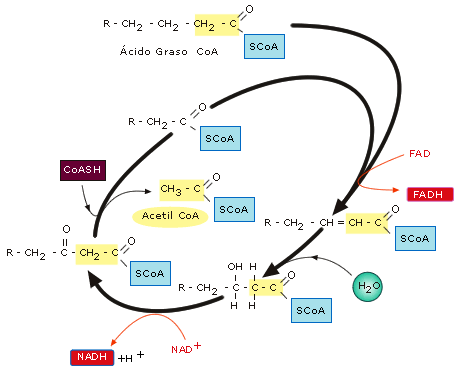
Figura
17: Esquema de la ß-oxidación de los ácidos grasos
Los transportadores
de electrones FADH2 y NADH liberan la energía que portan a lo
largo de la cadena respiratoria mitocondrial, mientras que el acetil-CoA
se incorpora al ciclo
de Krebs.
Al final, cada molécula de ácido graso puede aportar un gran número de
moléculas de ATP, más o menos dependiendo de su tamaño molecular y de
que sea saturado o no (por ejemplo, un ác. esteárico puede suministrar
hasta 146 moléculas de ATP).
3.7. El catabolismo de las proteínas
Las
proteínas, al igual que los azúcares y las grasas, también se pueden
descomponer para suministrar energía, pero no es ésta su principal función
en el organismo.
Los aminoácidos resultantes de la descomposición de las proteínas se
pueden degradar en dos fases: primero se eliminan los grupos amino y,
posteriormente, se oxida el "esqueleto carbonado" que quede.
Para eliminar los grupos amino son necesarios un proceso de transaminación
seguido de una desaminación,
formándose al final NH3. El amoníaco es una sustancia tóxica
que muchos animales, como los mamíferos, lo empaquetan en forma de urea
para su eliminación posterior a través de la orina.
El problema de los residuos nitrogenados, unido la valor de los aminoácidos
como "sillares estructurales" para los procesos de síntesis o
anabólicos, hace que las células sólo "quemen" las proteínas
cuando se ingieren en exceso o cuando, por el contrario, se agotan las
reservas de azúcares y lípidos, en situaciones de inanición.
4. Esquema general
del metabolismo
En el conjunto del metabolismo destacan dos procesos antagónicos en sus
productos iniciales y finales: la fotosíntesis y la respiración celular.
En ambos procesos ocurren intercambios energéticos y tienen lugar
fosforilaciones. También en ambos ocurren sendos ciclos de reacciones químicas
trascendentales: en la fotosíntesis, el ciclo de Calvin, en el que se
forma la primera materia orgánica; y en la respiración celular, el ciclo
de Krebs, en el que se descompone finalmente la materia orgánica.
Enlazando esos ciclos se encuentran tres intermediarios metabólicos básicos:
gliceraldehído 3-fosfato, piruvato y acetil-CoA.
Al final, en una célula que hipotéticamente pudiera realizar todos los
procesos metabólicos (sería autótrofa) entra O2, CO2,
H2O y luz; y sale O2, CO2, H2O
y ATP.
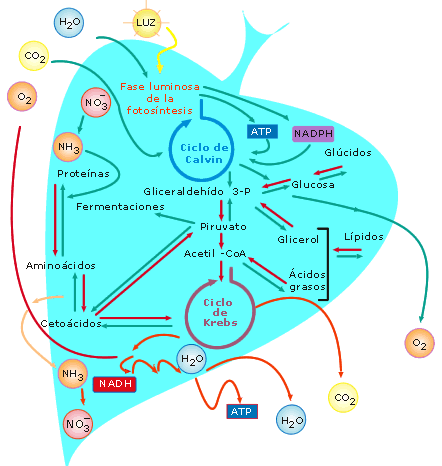
Figura
18: Esquema general del metabolismo, en una célula hipotética
que pudiera realizar todos los procesos metabólicos
básicos
 |