Portada.Historia.Tecnología.Sociedad.Enlaces
Plantas
y animales
Tanto los seres autótrofos (fotosintetizadores)
como los heterótrofos (descomponedores y animales) utilizan
el trifosfato de adenosina para intercambiar energía. Las
arquebacterias son organismos unicelulares a los que antes se clasificaba
como bacterias y que reducen compuestos inorgánicos sencillos. Las
bacterias tienen muchos taxones en ambos grupos tróficos
y desempeñan un papel imprescindible en los ciclos de la biosfera
que mantienen los flujos de nutrientes esenciales para la fotosíntesis.
Las plantas C3 y C4 son predominantemente autótrofas (los
cactus fijan el carbono por otro camino fotosintético). Los bosques
y las praderas son los biomas terrestres más importantes de
la biosfera con una biomasa que supera ampliamente la del fitoplancton
marino.
La energía necesaria para el metabolismo heterótrofo
procede, directa o indirectamente, de la digestión de cualquier tejido
de fitomasa disponible. Los herbívoros especializados pueden
digerir incluso la celulosa y la lignina que son sustancias abundantes pero
resistentes. Los carnívoros, aunque disponen de una menor
cantidad de biomasa para su alimentación, poseen una dieta más
nutritiva y de fácil digestión. Diferencias en el clima, la
producción vegetal y la biodiversidad heterótrofa dan lugar
a cadenas alimentarias que van desde simples enlaces a complicadas
redes.
Los animales homeotermos, pájaros y mamíferos que
regulan la temperatura de su cuerpo, poseen una incomparable adaptabilidad
ambiental, mientras que los poiquilotermos, organismos cuya temperatura
corporal es la del ambiente, son mucho más abundantes tanto en número
de especies como en número de individuos. Las diferencias entre estos
dos modos de existencia heterótrofa se manifiesta en los costes energéticos
de la reproducción, desarrollo y locomoción. Aunque
entre los poiquilotermos hay muchos y muy buenos nadadores, algunos
buenos corredores y también voladores, los mejores en natación
y, con mayor diferencia aún, en la carrera, salto y vuelo
son animales homeotermos.
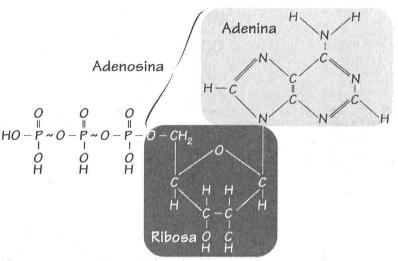
El ATP está formado por adenina, ribosa
y un terminal trifosfato.
Trifosfato de adenosina
En
bacterias, vegetales y animales, el
trifosfato de adenosina
(
ATP) es el enlace clave entre el catabolismo celular (degradación
de los nutrientes) de un lado y, por otra parte, el anabolismo (biosíntesis
de compuestos complejos), la locomoción (contracción muscular)
y el transporte activo de los metabolitos.
Este fosfato orgánico se forma durante el proceso de oxidación
de la glucosa y es la unidad de intercambio energético celular, porque
tiene una energía libre de hidrólisis intermedia (-31 kJ/mol).
Al inicio de la glicólisis cede un grupo fosfato para producir glucosa-6-fosfato
(con una energía libre de -13,8 kJ/mol) y puede ser fácilmente
formado, en una reacción posterior y mucho más exógena
(-49,3 kJ/mol), a partir del ácido 1,3-difosfoglicérico.
Durante el primer paso de estas reacciones catalizadas con enzimas,
la molécula de glucosa o de glucógeno se rompe para dar ácido
pirúvico. En un medio anaeróbico este proceso acaba en ácido
láctico (causando agujetas en los músculos) o en
etanol
y CO
2 (fermentación bacteriana). En presencia de oxígeno
se desarrolla el ciclo del ácido tricarboxílico, durante el
cual se transforman varios compuestos en CO
2, se trasfieren los
electrones liberados, se reduce oxígeno a agua y se producen grandes
cantidades de
ATP.
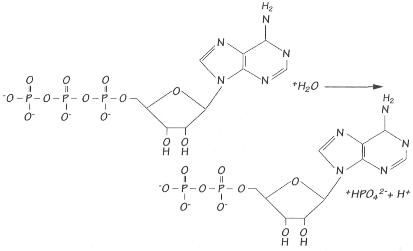
Cuando el ATP se hidroliza con agua para dar
adenosín-difosfato y un ion fosfato, se libera una gran cantidad
de energía.
En las células procariotas, la máxima
energía ganada en esta compleja secuencia es de 38 moles de ATP
por cada mol de glucosa, con una ganancia neta de energía libre
de 2,8 MJ y, dado que en cada transformación de ATP en difosfato
de adenosina se liberan 31 kJ/mol, la eficiencia total del proceso es de
un 40 por ciento. En las células procariotas, donde se llegan a liberar
hasta 50 kJ/mol en la transformación del ATP, la eficiencia
total puede superar el 60 por ciento.
La oxidación de los ácidos grasos alcanza una eficiencia
parecida, pero estos compuestos poseen una mayor densidad energética
que la glucosa. La máxima eficiencia en la transformación de
glucosa en ácido láctico es sólo de un 31 por ciento,
y únicamente algunos invertebrados que viven en ambientes poco oxigenados
utilizan habitualmente este proceso.
El adjetivo adecuado para describir la intensidad de generación
de ATP es apabullante. Los seres humanos sintetizamos y descomponemos
más de tres gramos de ATP por cada gramo de materia seca
de nuestro cuerpo, lo que significa que producimos y usamos diariamente
una cantidad de fosfato superior a la masa de nuestro propio cuerpo. Pero
esto no es nada comparado con la respiración de la bacteria Azotobacter,
una especie frecuente en los suelos y que forma amoníaco a partir
del nitrógeno atmosférico, que produce siete kilogramos
de ATP por cada gramo de materia seca de la que está formada.
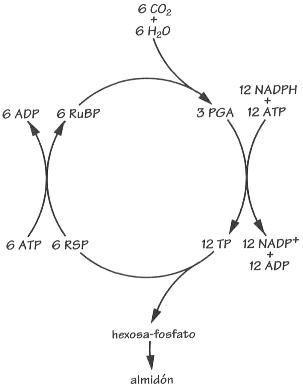
En el ciclo de la fotosíntesis el paso
de ATP a ADP, la reacción de reducción más importante
de la biosfera.
Estas intensidades de metabolismo heterótrofo
tan altas implican que los seres vivos desarrollan una potencia por unidad
de masa superior a la del Sol. La enorme masa solar (1,99 x 1033
g) y su inmensa luminosidad (3,9 x 1026 W) dan lugar a una intensidad
de 200 nW por gramo de masa solar, mientras que el metabolismo diario
de los niños (en promedio unos 3 mW/g de peso corporal) es 15.000
veces mayor y la respiración de las bacterias, que llega a
los 100 W/g, es 500 millones de veces superior a la del Sol. Por tanto,
aunque la magnitud de los flujos de energía de las estrellas es asombrosa,
la energía asociada con el ATP en los organismos heterótrofos
no tiene rival en cuanto a la intensidad de la conversión de energía
por unidad de masa.
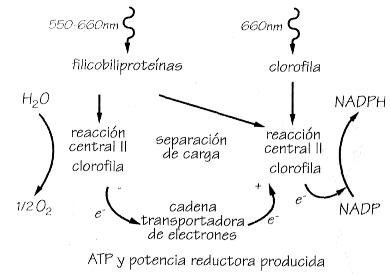
Esquema de la fotosíntesis en las bacterias
aeróbica y anaeróbica.
Ciclo anaeróbico.
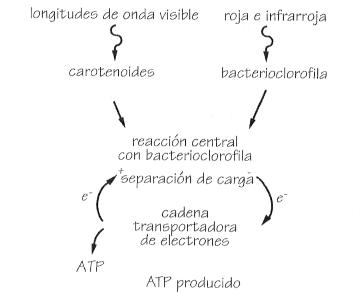 Ciclo aeróbico.
Ciclo aeróbico.
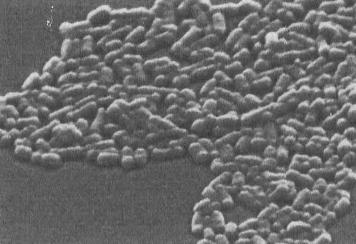
Grupo de bacterias Escherichia coli con forma de bastón,
que es un patógeno común humano.
Arquebacterias y bacterias
Estudios genéticos recientes iniciados por Carl Woese, y basados
en amplificaciones y clonaciones de secuencias de ARN, han conducido a una
nueva clasificación de los organismos vivos, asignando un nuevo reino
a las Arquebacterias. Vistos al microscopio, estos organismos unicelulares
no se distinguen de las bacterias, pero son genéticamente muy diferentes
de ellas, pareciéndose más a los eucuriotas, que constituyen
el tercer grupo de seres vivos en el que se incluyen los animales y vegetales.
Las arquebacterias actuales son descendientes de los organismos
más antiguos de la biosfera. El origen de las bacterias metanogénicas,
reduc-toras de CO2 y de otras formas simples de carbono (formiato,
metanol o acetato), así como las archaeons reductoras de sulfato,
se remontan a 4.000 millones de años. Muchos de estos organismos anaerobios
se desarrollan en ambientes extremos, terrestres y marinos —Methanococus
sobreviven en los fondos marinos bajo presiones superiores a las doscientas
atmósferas; Pyrodictium viven en las fuentes hidrotermales
profundas a temperaturas de entre 80 y 110°C y Archaeoglobus habitan
en pozos petrolíferos profundos— mientras que otros son frecuentes
en las tierras de cultivo.
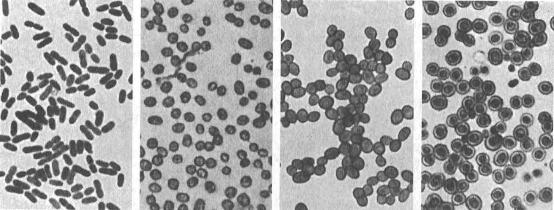
Estados de crecimiento de Azotobacter
vinelandii procedente del libro clásico Microbiologie du sol
de Winogradsky.
Todas las formas superiores de vida dependen de las
bacterias y de las arquebacterias, ya que ambas, a través
de los grandes ciclos bioquímicos, hacen circular los elementos vitales
tales como carbono, nitrógeno y azufre. No es sorprendente, pues,
que sean, con mucho, los organismos más abundantes en la biosfera y
que tengan un metabolismo muy adaptable: pueden ser anaerobios y aerobios,
así como autótrofos y heterótrofos fermentadores o respiradores.
Las primeras bacterias fueron quimioautótrofas, es decir, sintetizaban
biomasa sin radiación solar a partir de compuestos inorgánicos,
fundamentalmente oxidando nitrógeno y compuestos sulfurados.
Una vez más, estos organismos resultan indispensables para los
dos ciclos bioquímicos básicos. En el ciclo del nitrógeno
intervienen tomando energía de la transformación de NH3
en NO2 (Nitrosomonas) y de NO2 en NO3
(Nitrobacterias). En el ciclo del azufre obtienen la energía
oxidando sulfuros, tiosulfatos y sulfitos para producir sulfatas. Por otro
lado, hay otras bacterias anaeróbicas especializadas que reducen sulfatos
para producir H2S o azufre elemental.
Las bacterias que obtienen su energía de la fotosíntesis
incluyen organismos que producen biomasa tanto de forma anaeróbica
como aeróbica. La primera fotosíntesis, descubierta
en los años setenta, se realiza sin oxígeno y la segunda, más
frecuente, se produce en presencia de oxígeno. Las sulfurobacterias
púrpuras y las verdes, que viven en lodos sin oxígeno y que
utilizan H2S ó H2 como donantes de electrones
para reducir el CO2 pertenecen al filo de organismos anaerobios.
Hay dos tipos de fotosíntesis aeróbica, la que realizan,
sobre todo, las bacterias en ambientes marinos y la propia de las cianobacterias.
 Filamentos de Lynbya aestuarii, una cianobacteria que
forma densas acumulaciones en aguas bentónicas.
Filamentos de Lynbya aestuarii, una cianobacteria que
forma densas acumulaciones en aguas bentónicas.
Estos últimos organismos, conocidos antes como
algas verdiazuladas, sintetizan la biomasa de forma muy parecida a como lo
hacen las auténticas algas o los vegetales superiores. Son abundantes
en casi cualquier ambiente acuático, forman una parte importante del
fitoplancton y producen entre el 5 y el 20 por ciento de la fotosíntesis
marina. La extraordinaria adaptabilidad de las cianobacterias se demuestra
de las tres maneras siguientes: en sustratos nutritivos muchas especies terminan
siendo heterótrofas; en ecosistemas acuáticos pueden cambiar
de
fotosíntesis con oxígeno a
fotosíntesis
anoxigenica (basada en oxidación sulfurosa), lo cual demuestra claramente
la continua evolución de las estratégicas tróficas;
y, por último, otras especies son simbióticas con algas, hongos,
vegetales superiores y animales.
Otros seres heterótrofos que respiran son: las bacterias asociadas
con enfermedades en vegetales, animales y seres humanos, las actinobacterias
y las pseudomonas. Estos últimos incluyen los organismos descomponedores
que, o bien están especializados en algunos compuestos, tal como las
Cellulomonas que se alimentan de polisacáridos, o viven de
cualquier sustrato con carbono como las omnipresentes y móviles
Pseudomonas. Las bacterias heterótrofas constituyen una parte
importante del plancton (10-40 por ciento) tanto en las aguas costeras como
en mar abierto. Entre los fermentos heterótrofos anaerobios se encuentran
los implicados en las enfermedades infecciosas que van desde los accesos
dentales hasta la endocarditis. Las bacterias del suelo realizan una serie
de funciones insustituibles en los ciclos del carbono,
nitrógeno
y azufre. El contenido bacteriano de los suelos varía entre menos
de 100 kg de bacterias por hectárea en terrenos pobres, hasta nueve
toneladas por hectárea de los cultivos de alfalfa, siete de las cuales
se encuentran en la capa donde se desarrollan las raíces.
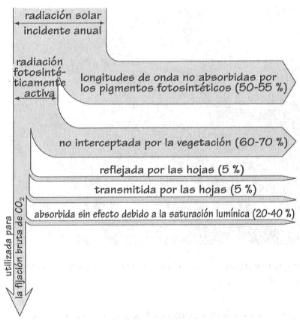 Distribución de la radiación solar que llega a
las plantas sintetizadoras.
Distribución de la radiación solar que llega a
las plantas sintetizadoras.
La mayor parte de las bacterias del suelo, generalmente
más del 70 por ciento, se encuentran en estado latente para poder
sobrevivir en ambientes pobres en nutrientes. Las bacterias del fitoplancton,
en aguas con pocos nutrientes, reaccionan a la inanición recurriendo
a una rápida fragmentación en pequeñísimas células.
Muchas bacterias no se comportan como células independientes, sino
como componentes de organismos multicelulares. Se separan en varias clases
de células diferentes, forman comunidades complejas que se mueven
siguiendo caminos marcados químicamente y se organizan en grupos depredadores.
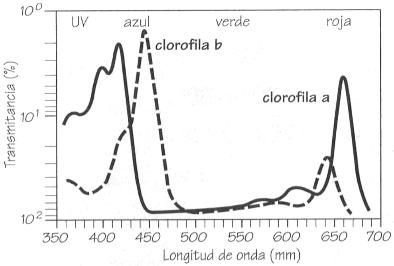
Absorción de luz por las diferentes clorofilas.
La fotosíntesis
La conocida ecuación básica que describe la reacción
endotérmica por la cual se sintetiza una molécula de glucosa
a partir de seis moléculas de CO2 y H2O, y 2,8
MJ de radiación solar, es una simplificadísima caja
negra. Una caja negra más realista sería la siguiente: 106
CO2 + 90 H2O + 16 NO3 + PO4 +
nutrientes minerales + 5,4 MJ de radiación = 3.258 g de protoplasma
( 106 C, 180 H, 46 O, 16 N, 1 P y 815 g de cenizas minerales) + 154 O2
+ 5,35 MJ de calor disipado. Sin macronutrientes ni micronutrientes no se
puede producir fitomasa, que está compuesta por los nutrientes básicos
necesarios para todos los seres heterótrofos: azúcares complejos,
ácidos grasos y proteínas.
Para entender la caja negra hay que empezar por destacar la acción
de unos pigmentos sensibles a la luz entre los cuales destacan las clorofilas.
Éstas absorben la luz en dos bandas estrechas, una entre 420 y 450
nm, y la otra entre 630 y 690 nm. Así, la energía necesaria
para la fotosíntesis sólo procede de la radiación
azul y roja a la que corresponde menos de la mitad de la energía total
de la insolación. Esta parte de la radiación fotosintéticamente
activa (RFA) no se utiliza en reducir CO2, sino en la regeneración
de compuestos consumidos durante la fijación del gas.
La síntesis de fitomasa en el ciclo reductor del fosfato pentosa
(RPP) —un proceso con varios pasos de carboxilación por catálisis
enzimática, reducción y regeneración— tiene que empezar
con la formación de trifosfato de adenosina (ATP) y nicotinamida
adenina dinucleótido fosfato (NADP), que son los dos compuestos que
suministran energía a todas las reacciones biosintéticas. La
síntesis de las tres moléculas de ATP y las dos de NADP que
se necesitan para reducir cada molécula de CO2 requiere
de diez cuantos de radiación solar con longitudes de onda próximas
al pico rojo de absorción de la clorofila (680 nm). El carbono procedente
del CO2, combinado con el hidrógeno del agua y con los
micronutrientes, dará lugar a nueva fitomasa que contiene 465 kJ/mol.
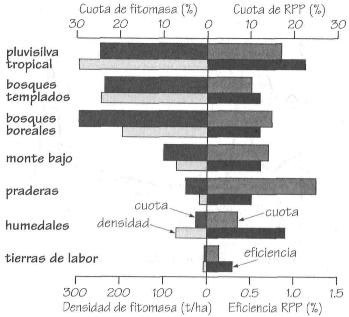
Cuotas y densidad de fitomasa, y cuotas y eficiencia de RPP, en los principales
ecosistemas.
La cantidad de energía de un cuanto de luz roja
es 2,92 x 10-19 J (el producto de la constante de Planck, 6,62
x 10-34 por la frecuencia de la luz, que, a su vez, es el cociente
entre la velocidad de la luz y la longitud de onda). Un einstein (definido
como un mol o el número de Avogadro, 6,02 x 1023) de fotones
rojos tiene una energía aproximadamente igual a 176 kJ. Suponiendo
que la RFA es aproximadamente el 45 por ciento de la luz directa, la eficiencia
global de la fotosíntesis es del 11 por ciento (456/[ 1.760/0,43]).
Esta eficiencia se reduce por lo menos en una décima parte más
si tenemos en cuenta la reflexión de luz en las hojas y la pérdida
que supone atravesar la cubierta vegetal.
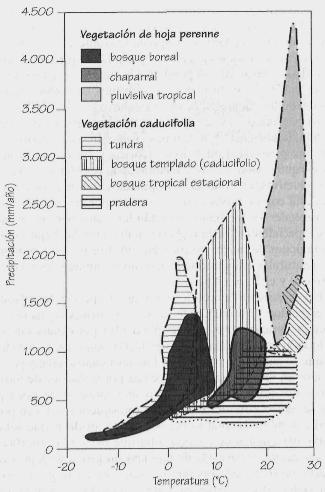
Los principales biotas delimitados por la precipitación y la temperatura.
Ninguna planta, sin embargo, se acerca siquiera a esta
eficiencia teórica porque parte de la luz absorbida por las clorofilas
(generalmente 20-25 por ciento) vuelve a ser emitida en forma de calor, debido
a que los pigmentos no pueden almacenar la luz y las reacciones enzimáticas
no se producen con suficiente velocidad como para utilizar completamente
el flujo de energía incidente. En la respiración se cede el
carbono fijado en el metabolismo de la planta y en el mantenimiento de las
estructuras que la soportan. Para cada especie la tasa con la que se pierde
carbono está determinada principalmente por el tipo de fotosíntesis.
Así, existen diferencias sustanciales entre las plantas C3
y C4. La respiración a escala de una comunidad
o ecosistema depende del estado de crecimiento, y varía entre menos
del 20 por ciento en plantas jóvenes, en rápido crecimiento,
hasta más del 90 por ciento en bosques maduros.
Con una pérdida del 25 por ciento para la reacción y otro
tanto para la respiración, la eficiencia fotosintética es ligeramente
superior al 5 por ciento. En este punto las estimaciones teóricas
y los valores reales coinciden, puesto que el valor máximo medido
de la fotosíntesis neta en plantas muy productivas,
en condiciones óptimas y durante cortos períodos de tiempo,
está comprendida entre el 4 y el 5 por ciento. Para la mayoría
de los vegetales los rendimientos están limitados por las carencias
de nutrientes, especialmente de nitrógeno y agua, o por las
bajas temperaturas en las regiones de mayor altura y latitud. Los mejores
rendimientos en sistemas naturales muy productivos como los humedales y los
cultivos están entre el 2 y el 3 por ciento.
En otros ecosistemas, como los pantanos tropicales templados y los bosques
templados, el rendimiento de la transformación es del 1,5 por ciento,
y en las praderas muy áridas sólo del 0,1 por ciento.
Globalmente, la producción anual es, al menos, de 100.000 millones
de toneladas de fitomasa, que se sintetizan con un rendimiento medio del
0,6 por ciento. La fotosíntesis en los océanos, muy
afectada por la escasez de nutrientes, es incluso menos eficiente. La productividad
media es de poco más de 3 M.J/m2 y se realiza con un rendimiento
fotosintético del 0,06 por ciento. La media ponderada total es 0,2
por ciento, es decir que solamente uno de cada 500 cuantos de energía
solar que llega a la superficie de la Tierra se transforma en energía
de biomasa en forma de tejido vegetal.
La mayor parte de esta energía se almacena en forma de azúcares
simples, en polisacáridos (especialmente celulosa) y en lignina.
Los lípidos, que contienen más energía, sólo
se encuentran en las semillas. Utilizando las mejores estimaciones y los
resultados de los almacenamientos en ecosistemas típicos, la fitomasa
total en los primeros años noventa se elevaba a un billón de
toneladas, que corresponde aproximadamente a 160 MJ/m2 de suelo.
La mayor parte de esta fitomasa se encuentra en los bosques, una parte
mucho menor en las praderas y menos del 0,5 por ciento en los cultivos agrícolas.
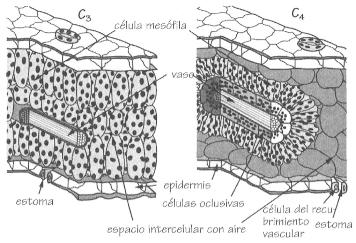 Secciones transversales de una hoja mostrando las diferencias
anatómicas entre las plantas C3 y C4.
Secciones transversales de una hoja mostrando las diferencias
anatómicas entre las plantas C3 y C4.
Plantas C3
y C4
No había pasado una década desde que Andrew Benson y Melvin
Calvin descubrieran la secuencia básica del ciclo RPP de la
fotosíntesis,
cuando Hal Hatch y Roger Slack demostraron que muchos
vegetales siguen
inicialmenle otro proceso de fijación del carbono. A diferencia de
lo que ocurre en el ciclo RPP, donde la reducción de CO
2
produce ácido fosfoglicérico, que es un compuesto con tres
átomos de carbono, hay
plantas que en sus células mesófilas
sintetizan primero oxaloacetato. Este ácido de cuatro carbonos es
reducido a malato, otro compuesto de cuatro carbonos, y transportado a los
cloroplastos de las células de la vaina que recubre el conjunto del
haz vascular, donde se transforma en CO
2 y piruvato. El piruvato
se reenvía a las células mesófilas y el CO
2
regenerado entra en el ciclo de Calvin incorporando su carbono en nueva biomasa
vegetal.
Debido a la reducción inicial de CO
2 en malato las
plantas
C4 poseen concentraciones internas muy bajas de este gas
—cerca de 10 ppm, comparadas con las 350 ppm del aire— y utilizan muy eficientemente
el agua. La presión de vapor de agua es mucho más alta en
el interior de las hojas que en la atmósfera, y ambas presiones son
dos órdenes de magnitud superiores a la diferencia de la presión
del CO
2 en el interior de la hoja y en la atmósfera. Por
tanto, la
fotosíntesis implica una gran pérdida de
agua hacia la atmósfera a cambio de la absorción de una pequeña
cantidad de CO
2.
| Características |
Plantas C3
|
Plantas C4
|
saturación luminosa
|
saturación a < 300 W/m2
|
no se satura
|
tasa máxima de fotosíntesis neta
(mg CO2/dm2 de área de hoja/h)
|
15-35
|
40-30
|
temperatura óptima para la fotosíntesis
(°C)
|
15-25
|
30-45
|
concentración de CO2 en la fotosíntesis
(ppm CO2)
|
30-70
|
0-10
|
fotorrespiracíón
|
alta
|
baja
|
tasa de transpiración (g H2O/g
de peso seco)
|
450-950
|
250-350
|
tasa de crecimiento (g peso seco/dm2
de área de hoja)
|
0,5-2
|
3-5
|
Principales diferencias entre las plantas C3
y C4.
En las plantas C4, mucho más
eficientes, este balance descompensado es de 450-600 moles de H2O
por cada mol de CO2, mientras que para las plantas C3
la relación se aproxima a 1.000 y puede llegar incluso hasta
4.000 moles de H2O.
Otra ventaja crítica de proceso C4 es que prácticamente
no hay fotorrespiración ni oxidación de biomasa vegetal sintetizada.
En los vegetales C3 todos los cloroplastos están
en las células mesófilas mientras que en los vegetales C4
se encuentran en las células de la vaina que cubre los tejidos vasculares.
Consecuentemente, en los vegetales C4 la ribulosa 1,5
difosfato carboxilasa —una enzima que puede utilizar tanto O2
como CO2 y que, por tanto, no sólo consume sino que también
produce el gas— está presente exclusivamente en las células
de la vaina donde los niveles de O2 son naturalmente bajos y
las posibilidades de oxidación despreciables.
La regeneración del piruvato requiere energía adicional que
es suministrada por el trifosfato de adenosina, pero dado que no
hay prácticamente fotorrespiración y que la carboxilasa piruvato
tiene una mayor afinidad por el CO2 que la carboxilasa ribulosa,
la ruta fotosintética de Hatch-Slack es bastante más eficiente
—hasta un 70 por ciento, promediando durante todo el Período de crecimiento—
que la fotosíntesis de las plantas C3.
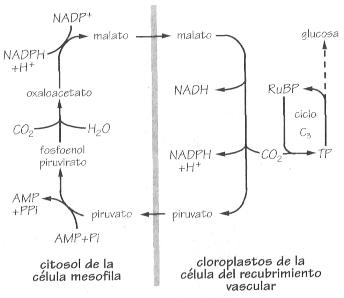
Secuencia básica del ciclo fotosintético
de las plantas C4.
La diferencia en el consumo de agua y en la productividad
primaria tiene unas implicaciones económicas enormes en los cultivos.
Así se explica que el maíz se haya convertido en el principal
cereal del mundo y que la producción de azúcar a
partir de la remolacha no pueda ser tan eficiente como a partir de la caña.
Muchas especies C4, tales como bigbluestem, bermudagrass,
paragrass y buffelgrass, contribuyen sustancialmente a la productividad
de las praderas. Desgraciadamente, los dos cereales preferidos, arroz
y trigo, así como todas las leguminosas, de alto contenido proteico,
son plantas C3.
Los plantas C4 tienen, además, otra ventaja climática.
Mientras que la fotosíntesis de los vegetales C3
se satura cuando la intensidad de la radiación solar alcanza
los 300 W/m2 y requiere una temperatura adecuada entre 15-25°C,
los vegetales C4 no se saturan por exceso de luz y funcionan
mejor a temperaturas comprendidas entre los 30 y 45°C, lo que hace que
los cultivos de estas últimas sean más competitivos en climas
soleados y cálidos.
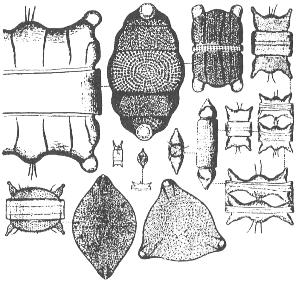
Grandes diatomeas del genero Biddulphia.
Fitoplancton
Aunque la mayor parte de la producción y almacenamiento de la fitomasa
terrestre se concentra en los grandes árboles de los
bosques,
en el océano los principales productores son los organismos que componen
el
fitoplancton, que son muy pequeños y flotan libres. Su tamaño
varía entre algo menos de 2 y 200
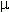
m de diámetro y están formados por cantidades variables de
bacterias y protoctistas eucarióticos. Las cianobacterias
cocoides son tan abundantes en algunas aguas oligotróficas que pueden
ser las responsables de la mayor parte de la producción de
fitoplancton.
Los protoctistas fotosintetizadores varían entre los más pequeños
flagelados pigmentados (como las criptomonas y crisofitos), hasta las diatomeas
y dinoflagelados, que son mayores (más de 10 mm) y generalmente dominantes.
Las diatomeas están formadas por células sin flagelos, con paredes
de silicio amorfo mezclado con otros compuestos orgánicos. Presentan
una sorprendente y amplia variedad de diseños, desde la que tienen
simetría central (las de forma radial son las dominantes en el océano),
a las pennadas (simetría lateral) y otras veces forman largas cadenas.
Las dinoflageladas se incluyen en un filo al que también pertenecen
numerosas especies no fotosintetizadoras.
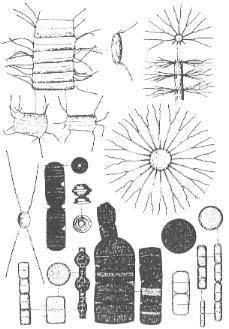 Diatomeas Chaetoceros delgados y Melosira segmentadas.
Diatomeas Chaetoceros delgados y Melosira segmentadas.
La productividad del fitoplancton está controlada
por la temperatura del agua y por la disponibilidad de radiación
solar y nutrientes. La temperatura rara vez es un factor decisivo, porque
muchas de las especies son muy adaptables y consiguen una productividad similar
en distintos ambientes. Aunque también es frecuente la adaptación
a diferentes condiciones de iluminación solar, tanto el volumen como
el contenido en clorofila de las diatomeas aumenta con la intensidad de luz.
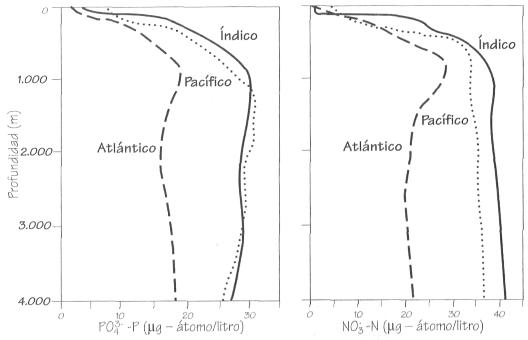
Distribución vertical de fosfato y nitrato
en los tres grandes océanos.
En el mar abierto la mayor limitación es la
cantidad de nutrientes disponible. Entre las carencias que más limitan
la producción de fitoplancton está la de nitrógeno,
el macronutriente más importante, de fósforo y de algunos otros
micronutrientes clave como el hierro y el silicio.
Entre los medios menos productivos de la Tierra se encuentran la
capa superficial y la capa inmediatamente inferior de los océanos.
En el mar abierto las concentraciones más altas de nutrientes se encuentran
entre los 500 y los 1.000 m, a bastante más profundidad que la zona
eufótica, capa en la que penetra la luz solar y que se extiende hasta
unos 100 m en las aguas más claras. El pequeñísimo tamaño
de los productores dominantes es una adaptación eficaz a la escasez
de nutrientes, ya que cuanto mayor sea el cociente entre la superficie y el
volumen, y más lento el hundimiento de las células de fitoplancton
en la capa eufótica, mayor es la tasa de absorción de nutrientes.
Cuando las corrientes elevan a la superficie las aguas frías y cargadas
de nutrientes, la producción de fitoplancton aumenta sustancialmente.
Las aguas costeras de Perú, California, noroeste y sudoeste de África,
y de la India occidental son ejemplos destacados de ascensión costera
de aguas frías. También se dan casos de ascensión mar
adentro en la mitad del Pacífico, cerca del ecuador y en las aguas
que rodean a la Antártida.
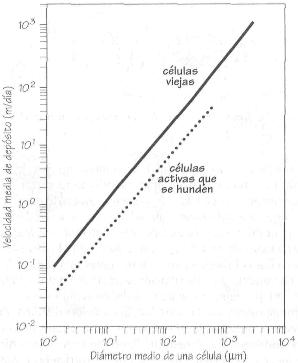
Ajustes lineales de la velocidad de depósito
del fitoplancton.
Otras zonas altamente productivas se encuentran en
las aguas poco profundas cercanas a la costa que están enriquecidas
por el aporte continental de nutrientes. Este enriquecimiento, con una proporción
N/P muy descompensada, es especialmente elevado en los estuarios adonde van
a parar grandes cantidades de aguas residuales y fertilizantes.
Las diferentes medidas de la productividad en las aguas oligotróficas
de los mares subtropicales y de las aguas eutróficas con corrientes
ascensionales varían entre menos de 50 g C/m2 y 1 g C/m2,
más de un orden de magnitud. Las estimaciones de la producción
global de fitoplancton están comprendidas entre 80.000 y 100.000
millones de toneladas, que representan entre dos tercios y cuatro quintos
de la fitomasa total terrestre. Contrasta con el resultado anterior el hecho
de que, dado el corto Período de vida del fitoplancton (1 -5
días), la fitomasa marina represente sólo una pequeña
fracción de todo el almacenamiento terrestre.
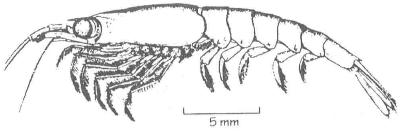
Ejemplar de zooplancton similar a la gamba,
común en el kril.
La distribución espacial del fitoplancton
muestra zonas delimitadas que se extienden a escala local y global. La exploración
desde los satélites es, con gran diferencia, la que permite detectar
con mayor precisión las concentraciones de clorofila y la que ha facilitado
obtener las pautas de la distribución del fitoplancton. En las
aguas que rodean la Antártida se observa claramente una distribución
asimétrica en dos bandas casi concéntricas. La mejor explicación
de esta distribución es que se deba a corrientes circumpolares y a
la abundancia de ácido silicílico. Pero las zonas con mayor
riqueza de fitoplancton se encuentran cerca de los continentes donde
los ríos arrastran abundantes nutrientes disueltos.
El fitoplancton es la base energética de las intricadas pirámides
tróficas. Las cadenas alimentarias en el océano
generalmente son redes complicadas. Una gran parte de la fitomasa disponible
no la consumen directamente los herbívoros, sino que primero
se almacena en depósitos de materia muerta que, transformada por bacterias,
se convertirá en alimento para otros seres heterótrofos. La
alta producción de fitoplancton puede alimentar grandes cantidades
de zoomasa. El kril, pequeños crustáceos parecidos a las quisquillas
que se alimentan de diatomeas, son los organismos más abundantes en
la superficie del mar; sus densas acumulaciones pueden contener hasta 1.000
millones de individuos y su producción anual de zoomasa quizá
llegue hasta los 1.300 millones de toneladas. Esta prodigiosa cantidad de
zoomasa sirve de alimento a focas, calamares y peces, así como a los
mayores carnívoros de la biosfera, las especies de ballenas
con barbas que se alimentan filtrando el agua.
"Energías" Vaclav Smil
Editorial Crítica S.L., Barcelona, 2001