|
Saccharomyces cerevisiae
El llevat de la
cervesa és una cèl·lula eucariotica
senzilla
Els
investigadors estan interessats en els eucariotes perquè nosaltres,
els humans, ho som. Les cèl·lules humanes, però,
són força complicades i resulta força difícil
treballar amb elles; si volem comprendre els elements fonamentals de
la biologia cel·lular dels eucariotes, sovint és més
eficient concentrar-se en altres espècies que, com E. Coli entre
les bactèries, són senzilles, robustes i es reprodueixen
ràpidament. L’espècie escollida de forma generalitzada
per aquest paper de model mínim eucariota ha estat el llevat Saccharomyces
cerevisiae(Fotografia 1) - el microorganisme utilitzat per a la
fabricació de cervesa, de vi i de pa-.
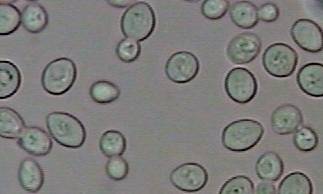
Beatriu Escudero
Fotografia
1.- Saccharomyces cerevisiae.
Saccharomyces cerevisiae és un petit fong unicel·lular.
De la mateixa manera que altres fongs, disposa d’una paret cel·lular
relativament rígida, és relativament immòbil, i
té mitocondris però no cloroplasts.
Quan
l’aliment és abundant, es reprodueix gairebé tan
ràpidament com un bacteri. El seu nucli tan sols conté una
quantitat d’ADN unes 2’5 vegades major que la de E. Coli,
per la qual cosa aquest llevat també és un bon subjecte
per als anàlisis genètics. Els estudis bioquímics
i genètics en llevats han estat crucials per a la comprensió de
molts dels mecanismes bàsics de les cèl·lules eucariotes,
incloent-hi el cicle de divisió cèl·lula (Veure
Annex I).

El
perquè de la seva importància biològica
i científica
El
llevat Saccharomyces cerevisiae (Fotografia 2) ha estat utilitzat
com a un sistema model per l´estudi d´altres
eucariotes superiors. Aquest microorganisme ha permès molts descobriments
en la investigació de la transcripció, traducció,
cicle cel.lular, transport de membrana,...
Sovint
ha estat triat degut a què:
- Aquest microorganisme creix amb facilitat.
- Té una gran quantitat de nucleòtids.
- Es té la complerta disponibilitat de la seva seqüència
genòmica.
- Pot ser manipulat
amb més simplicitat
que qualsevol altre eucariota.
És per totes aquestes raons que els gens dels mamífers
són introduïts rutinàriament en aquesta cèl·lula
per investigar la seva funció. A més, hi ha cents d´investigadors
en tot el món que usen aquest microorganisme activament.
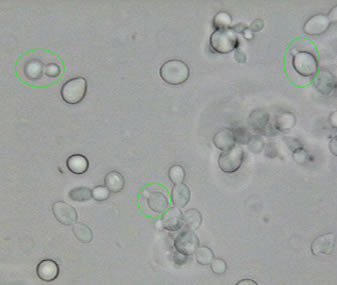
Beatriu Escudero
Fotografia
2.- Saccharomyces cerevisiae observat
al
microscopi òptic
reproduint-se asexualment
(gemmació)
Per
a conèixer millor
a aquest llevat ha estat necessari:
- Aïllar mutants.
- Classificar aquests mutants
en grups complementaris.
- Caracteritzar el seu fenotip.
- Clonar
els seus gens corresponents en genoteques de plàsmids.
Les
soques de Saccharomyces cerevisiae poden
existir en estat haploide o diploide. Hi ha dos tipus de cèl·lules
haploides:
#
Les “a” que quan s´aparellin donaran “Mat a”.
#
Les “α” que quan s´aparellin donaran “Mat α”.
Les
soques “a” i “alfa” poden aparellar-se donant
lloc a una soca diploide: “Mat a /α”.
“Mat a /α” és estable sota moltes
condicions i quan exposem aquesta soca sota unes altres condicions (concretament
un medi pobre, en una placa de Petri amb medi d’esporul·lació)
aquesta cèl·lula pot entrar en meiosi i donar quatre progenitors
haploides.
Aplicacions
Quan es va proposar seqüenciar
el genoma de Saccharomyces
cerevisiae sencer, molts investigadors es van qüestionar
la factibilitat i el valor del projecte. Pocs anys després,
la seqüenciació es va completar i actualment hi ha clares
evidències per demostrar que el projecte genoma era inapreciable
degut a què ha tingut moltíssimes aplicacions. Algunes
de les quals són:
Aquest microorganisme
s'ha utilitzat com a sistema model pel relatiu baix nombre de
gens que té el seu genoma, la extensió de
la similitud entre els seus gens i els d'eucariotes superiors i el
tractament dels seus sistemes experimentals.
L'objectiu
dels anàlisis genòmics comparatius és:
- Identificar gens
homòlegs funcionals, és
a dir, gens de llevats relacionats
amb gens d'altres organismes.
- Identificar gens
heteròlegs per poder fer estudis d'anàlisis
funcionals d'aquest microorganisme.
La utilitat d'aquest llevat
com un sistema per a entendre les funcions dels gens en eucariotes
superiors es basa en comparar les seqüències
dels gens de S.cerevisiae amb seqüències d'altres
organismes. Comparant totes les seqüències de proteïnes
conegudes en llevats amb les seqüències de mamífers,
a la base de dades del Genbank, es veu que aproximadament el 31% de gens
de llevats tenen homologia amb els gens de mamífers. Segurament és
una subestima del grau de conservació real, hi ha moltes seqüències
excloses que encara no es coneixen.
S'ha vist que els gens homòlegs entre llevats i
humans codifiquen per proteïnes involucrades en les principals vies
metabòliques i els gens que no tenen homologia codifiquen per:
receptors, components de la sang, components del sistema inmunitari i
per gens que no tenen similitud amb cap gen de funció coneguda.
A
l'hora d'interpretar aquests anàlisis comparatius s'ha de
tenir en compte que les seqüències homòlogues
no sempre indiquen conservació funcional. La conservació funcional
ve indicada per la homologia de la seqüència però no
es pot donar per suposat sense estudis addicionals. Una aproximació que
sovint es fa per determinar si es tracta d'un gen conservat o no, és
establir que la falta de funció d'un gen mutat en un llevat
pugui ser complementada per un gen homòleg d'un altre organisme.
Per tant, per fer estudis de conservació funcional entre llevats
i humans necessitem tenir una col·lecció complerta
de mutants delecionats de llevats i posseir la seqüència complerta
de cDNA per a cada gen humà.
-
Una altra aplicació ha estat: Estudi
de l'escala evolutiva: filogènia.
Es basa en l'estudi de famílies
de gens, per exemple, si una via metabòlica està present
en un organisme, aleshores, tots els gens requerits per aquesta funció estaran
presents. A més, si un gen es troba a un operó ha d'estar
involucrat en alguna funció: regulació, una via accessòria,...
La filogènia de famílies de gens ens pot
ajudar a conèixer les funcions ancestrals i les arrels del metabolisme
originari. Així, fent anàlisis comparatius es pot deduir
quins són els gens mínims requerits per a què una
cèl·lula funcioni, així com la reconstrucció d'una
cèl·lula mínima possiblement igual a la que va originar
la vida.
La seqüenciació d'entitats de
cromosomes permet també explorar la cursa de l'evolució predient
la força i les limitacions del genoma i comprendre el poder dels
microorganismes patògens per a fer laguerra.
Predir xarxes de regulació:
Analitzar
diferents genomes, ens ha permès saber els llocs dels
promotors, el lloc d'unió a proteïnes,...tot això és
important per tal de poder millorar la producció de determinats
metabolits microbians.
-
Saber com influeix la organització gènica
en el fenotip:
Comparant diferents organismes s'ha vist per exemple,
que hi ha fenotips diferents sense que hi hagi alteracions de seqüències
de gens ni guanys ni pèrdues de gens.
El genoma no és una estructura estàtica
ni físicament ni temporalment ni geneticament. És per això que
l'anàlisi de la seqüència genòmica ens pot
ajudar a identificar com actua la restricció sobre el genoma i
quin grau de fluïdesa és normal o tolerable.
- Estudi de l'ecologia microbiana:
L'estudi de la seqüenciació genòmica
també ens pot ajudar a identificar les eines que les cèl·lules
utilitzen per a fer front al món exterior i a predir com una cèl·lula
respon en un consorci d'altres cèl·lules, és a dir,
com és la interacció entre les diferents cèl·lules.
-
Aplicacions de Saccharomyces cerevisiae en
el descobriment de drogues (fàrmacs):
Aquest llevat és molt important sobretot en el
descobriment d'antifúngics. Les infeccions fúngiques en
humans afecten sovint a la pell o a les ungles produint malalties superficials
que poden arribar a ser sistèmiques. L'augment de pacients inmunodeprimits
ha fet que augmentin aquestes infeccions sistèmiques, juntament
amb la falta d'agents terapèutics. Això ha fet que hi hagi
una urgent necessitat d'identificar noves drogues per part de moltes
companyies farmacèutiques.
L'objectiu és identificar un agent que sigui actiu
en un ampli espectre de fongs patògens a través d'un mecanisme
fungicida. La molècula diana a la que vagi dirigit a aquest agent
ha d'estar present en l'espectre de gens patògens però no
en humans. A més, també ha de codificar per a una funció essencial
per a la viabilitat de la cèl·lula.
Probablement,
moltes de les funcions essencials pel creixement de S.cerevisiae també ho serà pel creixement d'altres espècies de
fongs. Per tant, s'haurà de fer anàlisis comparatius genòmics
per tal d'identificar gens essencials en aquest llevat que siguin conservats
en altres espècies de fongs i absents en humans (gens heteròlegs).
L'habilitat per expressar seqüències heteròlogues
codificades en aquests organismes ens proporcionar el poder desenvolupar
de forma ràpida i efectiva screenings per a nous agents terapèutics.
Per això fa falta que les seqüències heteròlogues
estiguin associades a un fenotip que es pugui mesurar fàcilment.
- Estudi de la interacció d'un
determinat lligam amb el seu receptor:
Actualment s'estan desenvolupant
sistemes per a identificar nous pèptids lligands de GPCRs (receptors acoplats a proteïnes
G). En Saccharomyces cerevisiae, els GPCRs i les proteïnes
G heterotrimèriques s'usen per a controlar el procés d'acoplament, és
a dir, les cèl·lules haploides de llevats expressen receptors
específics que s'uneixen a feromones secretades per cèl·lules
del tipus contrari d'aparellament. L'activació de receptors receptors
per la unió de lligands estimula una senyal de la via de transducció provocant
canvis en l'expressió de gens i en la seva morfologia que són
necessaris pel procés d'acoplament.
Plasmidis
L’ADN més utilitzat als experiments de transformació és
una petita molècula circular anomenada plasmidi. Aquests ADN circulars,
els plasmidis del Saccharomyces cerevisiae, són vectors "shuttle" que
poden introduir-se amb facilitat tant en Escherichia coli com
en aquest llevat. És per això que aquests plàsmids
porten orígens de replicació i marcadors selectius per
a mantenir-se a dita bactèria i al llevat.
Gràcies a què aquests
plàsmids
poden introduir-se amb facilitat dins d´aquests microorganismes,
alguns gens que corresponen a un fenotip mutant poden ser aïllats
per complementació amb una genoteca de plasmidis. A més,
utilitzant-los per ells mateixos o amb combinació amb un promotor
induïble podem sobreexpressar gens homòlegs i heteròlegs.
La integració del DNA plasmídic transformat dins del genoma
es fa per recombinació homòloga.
Genoma de Saccharomyces cerevisiae
Es va acabar de seqüenciar
a l´Abril
del 1996. Aquest fet va ser important per dues raons:
- Perquè va ser el primer genoma eucariota seqüenciat.
- Per la forma en què va
ser obtingut, ja que ho va fer un consorci de laboratoris
de tres continents.
El
seu genoma té 16 cromosomes (figura 1) que tenen un rang de
llargada de 200 a 2200 quilobases (Kb) i la seva seqüència
genòmica
total dipositada a les bases de dades és de 12,1 Megabases (Mb).
Aparentment el seu genoma és ric en informació:
- Té 70% de gens dedicats a codificar
seqüències.
- En total s'han identificat
6215 seqüències potencialment
codificadores de proteïnes.
- El promig de la mida
d'un marc obert de lectura (ORF) és
de 1450 parells de bases.
Els introns( o fragments d’ADN que no s’expressen) no
són comuns i els que existeixen són curts, el més
llarg fa 1 Kb i primàriament estan a l’extrem 5' dels
gens.
Quan
la seqüència va ser publicada, sol s'havia descobert
el 30% de la seqüència codificadora per tècniques
convencionals. Per al restant 70% hi havia alguna infomació funcional
que podia ser deduïda per la homologia amb altres gens de funció coneguda.
No obstant, un terç de gens ni tenien una seqüència
homòloga amb els gens de les bases de dades ni amb altres ORFs
hipotètics o no caracteritzats.
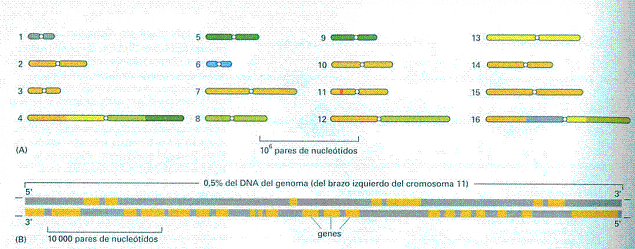
Figura 1.-
Cromosomes de Saccharomyces cerevisiae
Degut
a què hi havia hagut tanta informació indetectada,
es van preguntar perquè tants ORFs no havien estat detectats:
- Es va pensar que
aquests ORFs podien no tenir cap informació biològica. Aquesta
hipòtesi va ser refusada quan van detectar la transcripció en
el 87% dels gens del genoma.
- L'explicació més probable a això és la
forma en què els previs experiments genètics van estar
dirigits: les primeres proves van ésser configurades per a identificar
gens que juguen un paper més important i van estar decantades
cap a mutants amb forts fenotips com per exemple els mutants letals
condicionals. En aquestes proves, les mutacions que van donar un fenotip
dèbil haurien estat excloses o indetectades.
També és possible que la majoria dels nous gens siguin
requerits per a un creixement sota condicions no encara reproduïbles
al laboratori.
El genoma de Saccharomyces cerevisiae té un elevat nivell
de seqüències redundants. L'anàlisi d'aquesta seqüència
indica que:
- Aproximadament el 25% dels seus
gens existeixen com a parells de gens homòlegs.
-
D'aquests, 376 parells de gens resideixen en unes regions del cromosoma
anomenades CHR (Cluster homology regions: agrupacions de regions
homòlogues).
Aquestes regions tenen 3 o més gens homòlegs en el mateix
ordre i orientació relativa. La llargada d'aquests CHRs varia
de 7.5 a 170 Kb i ocupa aproximadament el 50% del genoma. La majoria
d'aquests CHRs han guanyat o perdut components com gens, introns, elements
transponibles o pseudogens.
S'ha
proposat que els CHRs van ser generats simultàniament en
un genoma d'àmplia duplicació fa 100 milions d'anys aproximadament.
A més de tot això,
també hi ha un gran
número de parells de gens homòlegs localitzats fora dels
CHRs, així com un número més petit de famílies
de gens.
Solament
els 2'5% de gens essencials s'han trobat en els CHRs. Es creu que els
homòlegs d'aquests gens essencials són o no funcionals
o han evolucionat cap a una funció diferent.
Un
dels primers passos per a entendre la funció d'un gen és
obtenir informació sobre aquest gen, és a dir, per a cada
gen es necessita caracteritzar:
-
La seva expressió genòmica.
-
L'expressió i la modificació de les proteïnes.
-
Les interaccions proteïna-proteïna.
-
Les conseqüències de la cèl·lula
al perdre aquest gen.
A
diferència de les seqüències genòmiques,
aquestes dades són dinàmiques i canvien en resposta als
factors interns i externs, a les condicions del medi i a la presència
de mutants.
El cicle vital de Saccharomyces cerevisiae inclou
fases haploides i diplomades
A
partir tant de cèl·lules eucariòtiques com procariòtiques
es poden obtenir poblacions genèticament homogenies. Per exemple,
a partir d’una sola cèl·lula vegetativa (cèl·lula
en creixement) o d’una sola espora, es poden obtenir cultius de
llevats i floridures. Els temes d’investigació especialment
ben estudiats amb aquests organismes han estat els mecanismes de control
cel·lular de la síntesi d’ADN, l’estructura
del cromosoma i la reordenació i replicació dels cromosomes.
Molts
espècies de llevats,
entre elles el Saccharomyces cerevisiae (o
llevat de panificació), creixen a la natura amb forma de cèl·lules
diploides individuals. Cada cèl·lula té, segons
la espècie, de 12 a 18 cromosomes.
Les
cèl·lules
del Saccharomyces cerevisiae es
reprodueixen asexualment mitjançant la gemmació. La gemma
comença a formar-se com una petita protuberància en una
regió eixamplada de la paret cel·lular anomenada “placa”.
La gemma va augmentant gradualment de tamany i finalment es separa de
la cèl·lula parental. La divisió del nucli en els
llevats no és tan complexa com a la majoria de les cèl·lules
eucariòtiques; no es realitza una mitosi vertadera. Els cromosomes
estan units a una estructura anomenada “cos polar del fus” -SPB
de l’anglès spindle pole body- . Quan
es repliquen els cromosomes, el cos polar del fus també ho fa
i l’estructura del fus separa els dos SPB, de manera que un d’ells
pot colocar-se a la gemma. El material cromosòmic
duplicat que està al nucli parental es reparteix així entre
les dues cèl·lules.
Beatriu Escudero
Fotografia 3.-Divisió cèl·lular
a Saccharomyces cerevisiae
Quan
una cèl·lula diploide troba condicions adverses (com
manca d’aliment), experimenta una forma de diferenciació per
a transformar-se en espora. Per a ser precisos, la cèl·lula
diploide inicialment experimenta divisions meiòtiques (veure l’Annex
I) produint quatre espores haploides. Si les condicions adverses desapareixen,
les espores es fusionen formant dues cèl·lules diploides;
en cas contrari, cada espora pot donar lloc a una colònia haploide
(Figura 3).
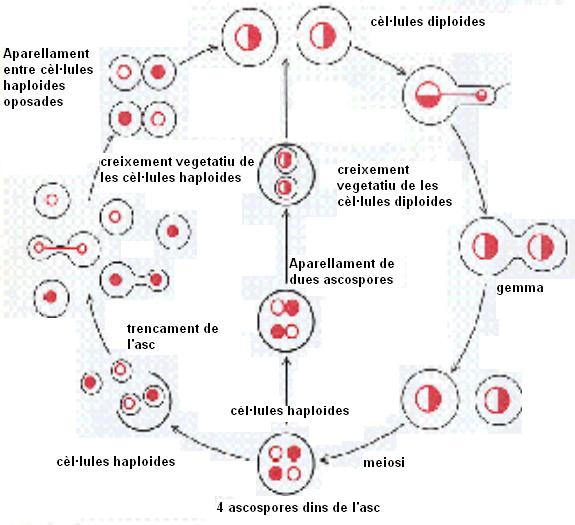
Figura
2.- Cicle biologic de Saccharomyces
cerevisiae
Les dues porcions haploides de cada
cèl·lula
diploide es diferencien en el tipus d’aparellament , degut
a que l’aparellament pot dur-se a terme solament entre components
haploides que poseeixen un tipus d’aparellament diferent. En
condicions de creixement favorables, les cèl·lules
diploides es reprodueixen asexualment per gemmació. Tanmateix,
quan a una cèl·lula li manquen els nutrient essencials,
sintetitza una paret exterior molt resistent, a partir de la qual
es forma unaasc. A seu interior , la cèl·lula
duu a terme una meiosi i els cromosomes es reparteixen independentment
segons el model mendelià, donant lloc a quatre ascospores haploides.
Si l’asc torna a un medi favorable,les parelles d’espores
poden fusionar-se entre elles per formar cèl·lules
diploides, que es desenvoluparan per gemmació com abans. D’altra
banda, cada ascospora pot romandre en estat haploide donant lloc
a una colonia haploide. Les cèl·lules haploides que
provinguin de diferents colonies (és a dir, cèl·lules
hapliodes amb diferents dotacions cromosòmiques) poden fusionar-se
(o ser fusionades al laboratori) formant cèl·lules
diploides.
A
la natura, la fusió de dues cèl·lules haploides
de llevat de diferents soques probablement passa poc sovint; quan
això es
produeix, les espècies augmenten el fons genètic disponible.
Al laboratori, la fusió intencionada de dues cèl·lules
haploides diferents es denomina “creuament genètic”.
Aquest procediment permet l’examen genètic de cada cèl·lula
parental.
La
fusió de determinades soques de cèl·lules haploides
pot produir-se gràcies a que les cèl·lules poden
actuar com “tipus d’aparellament”. Solament es poden
fusionar cèl·lules que difereixin en el seu tipus d’aparellament
per donar lloc a cèl·lules diploides.
La
expressió del tipus reproductor en llevats inclou una sofisticada
xarxa de gens que interaccionen entre sí i que està governada
per l’acció d’altres gens reguladors. Les cèl·lules
haploides del llevat presenten els tipus reproductors “a” o “α”. Cada
tipus reproductorprodueix un oligopèptid (un factor “a” o “α” respectivament)
que s’uneix a les cèl·lules del tipus oposat i atura
el seu creixement. Així, cada cèl·lula tendeix a
agregar-se amb una cèl·lula del tipus oposat -“shmoo
a” i “shmoo α” de manera que un shmoo es diferencia
amb força claredat d’una cèl·lula normal del
mateix llevat (Fotografia 4) i la cèl·lula diploide resultant
s’anomena a/α. Els diploides a/α poden
experimentar meiosi i esporulació per a produir de nou cèl·lules “a” i “α”.
El tipus cel·lular d’una cèl·lula haploide
està determinat per la seqüència, “a” o “α”,
que es troba al locus MAT; al MAT es produeixen dos parells de ARNm diferents,
un parell específic d’“a” i l’altre
d’“α”. Les copies emmagatzemades
d’ADN dels tipus “a” i “α” es
troben en dos loci silenciosos (és a dir, que no s’expressen),
HML i HMR, (llocs a l’esquerra i a la dreta, respectivament, de
MAT al cromosoma III). Normalment no es transcriu cap ARN a partir de
les seqüències de HML i HMR.

Beatriu Escudero
Beatriu Escudero
Fotografia
4.- Dos Shmoos (a les dues imatges superiors) i cèl·lules
en
estat normal de Saccharomyces cerevisiae.
Quan
s’aparellen (Figura 2) haploides genèticament diferents
per donar lloc a cèl·lules diploides que, al seu torn,
se’ls permet experimentar meiosi i segregació a noves espores
haploides, es pot estudiar la varietat de cromosomes resultants de la
primera divisió meiòtica. Aquests experiments han permès
establir “grups de segregació” (grups de gens que
es localitzen al mateix cromosoma). Pot detectar-se un grup de segregació per
l’alta freqüència de reaparició de combinacions
d’al·lels parentals a les cèl·lules de la
progenie (un al·lel és una de les diverses formes homòlogues
que pot adoptar un gen determinat). La localització dels gens és
possible gràciesa que durant la meiosi es produeixen entrecreuaments.
S’han aïllat moltes soques mutants de cèl·lules
de llevats i localitzat les seves mutacions al cromosoma. Avui en dia
existeixen cents de mutants nutricionals, és a dir, cèl·lules
que requereixen nutrients que una cèl·lula normal no els
requeriria, així com soques que es troben bloquejades en diverses
fases del seu cicle cel·lular normal.
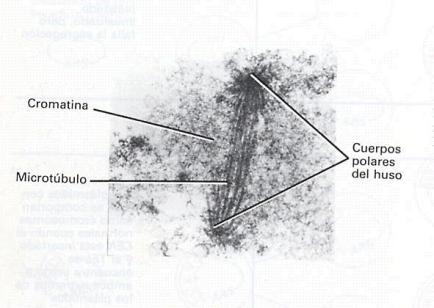
Figura
3.-Micrografia electrònica de
cossos polars del fus del llevat amb els seus microtúbuls units.
La cèl·lula de llevat ha estat fixada i seleccionada,
de manera que alguns dels microtúbuls no es poden veure. Tanmateix,
a través de l’estudi de seccions seriades, s’ha
vist que el nombre de microtúbuls és aproximadament igual
al nombre de cromosomes.
Com
s’ha esmentat anteriorment, la majoria dels llevats creixen
a la natura com organismes diploides però, com alguns fongs, formen
espores haploides com a resposta a certs canvis al medi ambient. En germinar,
les espores haploides formen cèl·lules de dos tipus (denominades “a” i “α” en Saccharomyces
cerevisiae de la manera com s’ha explicat abans); dues cèl·lules
de tipus reproductor oposat es fusionen regularment per a formar diploides.
Els llevats comparteixen un pol gènic gran i variat que els permet
adaptar-se a situacions ambientals i ecològiques diverses, tot
això degut a que la fusió de les cèl·lules
haploides pot realitzar-se entre cèl·lules de diferents
dotacions gèniques.
El
procés complet d’esporul·lació es realitza
aproximadament en 24 hores, mentre que la divisió d’una
cèl·lula vegetativa es completa en solament 2 hores. El
procés meiòtic que forma espermatozoides i oòcits
als organismes superiors té algunes semblances amb la esporul·lació dels
llevats diploides. Després de la duplicació de l’ADN,
cada cèl·lula premeiòtica es 4n (és
a dir, té 4 copies de cada cromosoma, dues maternes i dues paternes).
Aleshores la divisió meiòtica redueix els 4n a
1n als espermatozoides, oòcits i espores de llevat.
Ambdues
divisions de cromosomes es produeixen, als llevats, dins d’una
membrana nuclear simple. Després de completar-se la segregació dels
cromosomes, la membrana nuclear original es torna a formar i envolta
a cada un dels quatre nuclis haploides nous de les espores.
Al
mateix temps, el material de la paret cel·lular envolta cada
nucli; finalment es formen quatre espores haploides madures. La esporul·lació dels
llevats presenta diversos avantatges per a la investigació enfront
a la producció d’espermatozoides i oòcits en organismes
multicel·lulars: en llevats poden obtenir-se mutants en els quals
la meiosi es bloqueja en diversos passos; també pot induir-se
la meiosi sincrònicament en gran quantitat de cèl·lules
de llevat diploides.
En
llevats, la iniciació provoca la formació d’espores
i la meiosi. Durant la esporul·lació gran part de l’ARN
i proteïnes preexistents es degraden (aproximadament el 50%) i es
formen nous ARNm i proteïnes. Els nous complexes multiproteics generats
abans i durant la meiosi inclouen el complex sinaptonèmic que
participa en la recombinació dels cromosomes.
S’han detectat als llevats
un nombrós grup de mutacions
que bloquegen el cicle de divisió de les cèl·lules
vegetatives. Els gens identificats per a aquestos mutants es denominen
gens CDC (cicle de divisió cel·lular). La majoria d’aquests
gens proporcionen les funcions necessàries a la meiosi i a la
mitosi per la qual cosa són necessaris tant per al creixement
cel·lular vegetatiu com per a ala esporul·lació.
A més, entre 50 i 100 gens codifiquen proteïnes que són
essencials per a la esporulació i per a la meiosi, però no
per a la mitosi ni per al creixement cel·lular vegetatiu. Aquests
gens inclouen els que codifiquen enzims que formen el material de la
paret de l’espora i els que codifiquen proteases que destrueixen
les proteïnes preexistents durant la formació de l’espora.
Mutacions en alguns d’aquests gens interrompen la iniciació de
la esporul·lació; altres bloquegen etapes discretes a la
ruta de la esporul·lació.
Moltes
proteïnes específiques de la esporulació encara
no s’han aïllat però les anàlisis genètiques
suggereixen que aquestes proteïnes es formen seqüencialment
durant el procés d’esporul·lació. L’anàlisi
molecular continuat de l’esporul·lació de llevats
proporcionarà informació que pot ser important per a la
comparació de processos comparables amb les cèl·lules
superiors.
|